
25.4 Seedless Vascular Plants
Learning Outcomes
- Identify the new traits that first appear in seedless vascular plants
- Discuss how each trait is important for adaptation to life on land
- Identify the classes of seedless tracheophytes
- Describe the life cycle of a fern
- Understand the importance of seedless plants to humans
Vascular Plants
The vascular plants are the dominant and most conspicuous group of land plants. More than 260,000 species of land plants represent more than 90 percent of Earth’s vegetation. Several evolutionary innovations explain their success and their ability to spread to all habitats.
Bryophytes may have been successful at the transition from an aquatic habitat to land, but they are still dependent on water for reproduction, and must absorb moisture and nutrients through the gametophyte surface. The lack of roots for absorbing water and minerals from the soil, as well as a lack of lignin-reinforced conducting cells, limit bryophytes to small sizes. Although they may survive in reasonably dry conditions, they cannot reproduce and expand their habitat range in the absence of water. Vascular plants, on the other hand, can achieve enormous heights, thus competing successfully for light. Photosynthetic organs become leaves, and pipe-like cells or vascular tissues transport water, minerals, and fixed carbon organic compounds throughout the organism.
Throughout plant evolution, there is a progressive increase in the dominance of the diploid sporophyte generation. In seedless vascular plants, the sporophyte is the dominant phase of the life cycle. The gametophyte is now less conspicuous, but still independent of the sporophyte. Seedless vascular plants still depend on water during fertilization, as the flagellated sperm must swim on a layer of moisture to reach the egg. This step in reproduction explains why ferns and their relatives are more abundant in damp environments.
Vascular Tissue: Xylem and Phloem

The first plant fossils that show the presence of vascular tissue date to about 430 million years ago. The simplest arrangement of conductive cells shows a pattern of xylem at the center surrounded by phloem. Xylem is the tissue responsible for the long-distance transport of water and nutrients. Xylem conductive cells incorporate the compound lignin into their walls, and are thus described as lignified. Lignin itself is a complex polymer: It is impermeable to water and confers mechanical strength on vascular tissue. With their rigid cell walls, the xylem cells provide support to the plant and allow it to achieve impressive heights. Tall plants have a selective advantage by being able to reach unfiltered sunlight and disperse their spores or seeds away from the parent plant, thus expanding the species’ range.
Phloem is the second type of vascular tissue; it transports sugars, proteins, and other solutes throughout the plant. Phloem cells are divided into sieve elements (conducting cells) and cells that support the sieve elements. Together, xylem and phloem tissues form the vascular system of plants (Figure 25.16).
Roots: Support for the Plant
Roots appeared later in evolution than vascular tissue. The development of an extensive network of roots represented a significant new feature of vascular plants. Thin rhizoids attached bryophytes to the substrate, but these rather flimsy filaments did not provide a strong anchor for the plant; nor did they absorb water and nutrients. In contrast, roots, with their prominent vascular tissue system, transfer water and minerals from the soil to the rest of the plant. The extensive network of roots penetrates deep into the soil to reach sources of water also stabilizes plants by acting as an anchor. The majority of roots establish a symbiotic relationship with fungi, forming mutualistic mycorrhizae, which benefit the plant by greatly increasing the surface area for absorption of water, soil minerals, and nutrients.
Leaves, Sporophylls, and Strobili
A third innovation marks the seedless vascular plants. Accompanying the development of vascular tissue, the appearance of true leaves improved photosynthetic efficiency. Leaves capture more sunlight with their increased surface area by employing more chloroplasts to trap light energy and convert it to chemical energy, which is then used to fix atmospheric carbon dioxide into carbohydrates. The carbohydrates are transported to the rest of the plant by the conductive cells of phloem tissue.
The existence of two types of leaf morphology—microphylls and megaphylls—suggests that leaves evolved independently in several groups of plants. Microphylls (“little leaves”) are small and have a simple vascular system. A single unbranched vein—a bundle of vascular tissue made of xylem and phloem—runs through the center of the leaf. Megaphylls most likely appeared independently several times during the course of evolution. Megaphylls are seen in ferns and more derived vascular plants.
In addition to photosynthesis, leaves play another role in the life of the plants. Pine cones, mature fronds of ferns, and flowers are all sporophylls—leaves that were modified structurally to bear sporangia. Strobili are cone-like structures that contain sporangia. They are prominent in conifers, where they are commonly known as pine cones.
Ferns and Other Seedless Vascular Plants
By the late Devonian period, plants had evolved vascular tissue, well-defined leaves, and root systems. With these advantages, plants increased in height and size. During the Carboniferous period (360 to 300 MYA), swamp forests of club mosses and horsetails covered most of the land. These forests gave rise to the extensive coal deposits that gave the Carboniferous its name. In seedless vascular plants, the diploid sporophyte became the dominant phase of the life cycle.
Water is still required as a medium of sperm transport during the fertilization of seedless vascular plants, and most favor a moist environment. Modern-day seedless vascular plants include club mosses, horsetails, ferns, and whisk ferns.
Phylum Lycopodiophyta: Club Mosses

The club mosses, or phylum Lycopodiophyta, are the earliest group of seedless vascular plants. They dominated the landscape of the Carboniferous, growing into tall trees and forming large swamp forests. Today’s club mosses are diminutive, evergreen plants consisting of a stem (which may be branched) and microphylls (Figure 25.17). The phylum Lycopodiophyta consists of close to 1,200 species, including the quillworts (Isoetales), the club mosses (Lycopodiales), and spike mosses (Selaginellales), none of which are true mosses or bryophytes.
Lycophytes follow the pattern of alternation of generations seen in the bryophytes, except that the sporophyte is the major stage of the life cycle. Some lycophytes, like the club moss Lycopodium, produce gametophytes that are independent of the sporophyte, developing underground or in other locations where they can form mycorrhizal associations with fungi. In many club mosses, the sporophyte gives rise to sporophylls arranged in strobili, cone-like structures that give the class its name. Sporangia develop within the chamber formed by each sporophyll.
Lycophytes can be homosporous (spores of the same size) or heterosporous (spores of different sizes). The spike moss Selaginella is a heterosporous lycophyte. The same strobilus will contain microsporangia, which produce spores that will develop into the male gametophyte, and megasporangia, which produce spores that will develop into the female gametophyte. Both gametophytes develop within the protective strobilus.
Phylum Monilophyta: Class Equisetopsida (Horsetails)
Horsetails, whisk ferns, and ferns belong to the phylum Monilophyta, with horsetails placed in the class Equisetopsida. The single genus Equisetum is the survivor of a large group of plants, known as Arthrophyta, which produced large trees and entire swamp forests in the Carboniferous. The plants are usually found in damp environments and marshes (Figure 25.18).

The stem of a horsetail is characterized by the presence of joints or nodes, hence the name Arthrophyta (arthro- = “joint”; -phyta = “plant”). Leaves and branches come out as whorls from the evenly spaced joints. The needle-shaped leaves do not contribute greatly to photosynthesis, the majority of which takes place in the green stem (Figure 25.19).

Silica collected by in the epidermal cells contributes to the stiffness of horsetail plants, but underground stems known as rhizomes anchor the plants to the ground. Modern-day horsetails are homosporous. The spores are attached to elaters—as we have seen, these are coiled threads that spring open in dry weather and casts the spores to a location distant from the parent plants. The spores then germinate to produce small bisexual gametophytes.
Phylum Monilophyta: Class Psilotopsida (Whisk Ferns)
While most ferns form large leaves and branching roots, the whisk ferns, class Psilotopsida, lack both roots and leaves, probably lost by reduction. Photosynthesis takes place in their green stems, which branch dichotomously. Small yellow knobs form at the tip of a branch or at branch nodes and contain the sporangia (Figure 25.20). Spores develop into gametophytes that are only a few millimeters across, but which produce both male and female gametangia. Whisk ferns were considered early pterophytes. However, recent comparative DNA analysis suggests that this group may have lost both vascular tissue and roots through evolution, and is more closely related to ferns.

Phylum Monilophyta: Class Polypodiopsida (True Ferns)

With their large fronds, the true ferns are perhaps the most readily recognizable seedless vascular plants. They are also considered to be the most advanced seedless vascular plants and display characteristics commonly observed in seed plants. More than 20,000 species of ferns live in environments ranging from the tropics to temperate forests. Although some species survive in dry environments, most ferns are restricted to moist, shaded places. Ferns made their appearance in the fossil record during the Devonian period (420 MYA) and expanded during the Carboniferous (360 to 300 MYA).
The dominant stage of the life cycle of a fern is the sporophyte, which consists of large compound leaves called fronds. Fronds may be either finely divided or broadly lobed. Fronds fulfill a double role; they are photosynthetic organs that also carry reproductive organs. The stem may be buried underground as a rhizome, from which adventitious roots grow to absorb water and nutrients from the soil; or, they may grow above ground as a trunk in tree ferns (Figure 25.21). Adventitious organs are those that grow in unusual places, such as roots growing from the side of a stem.
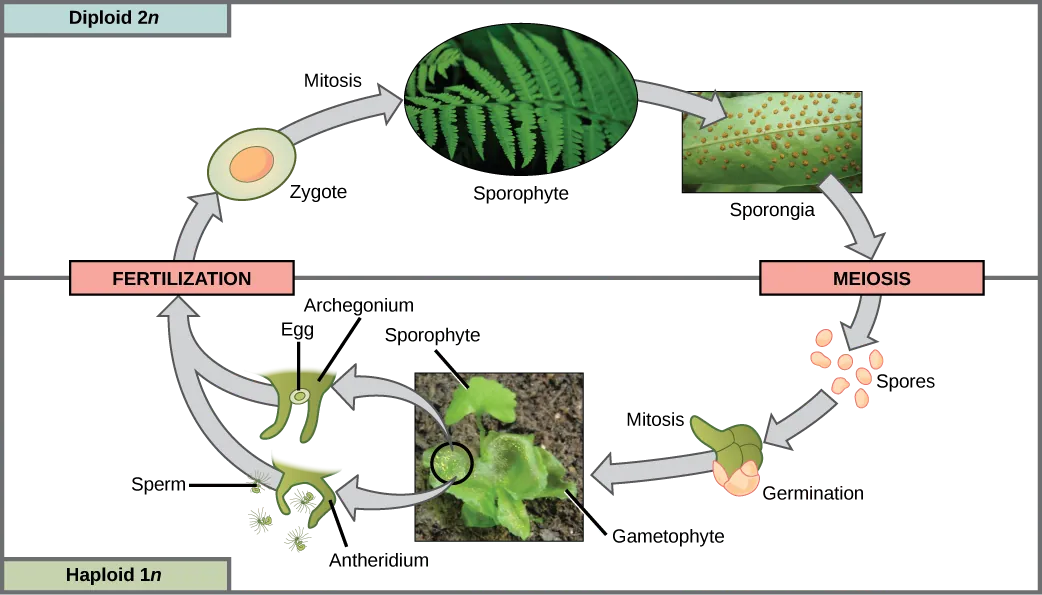
On the underside of each mature fern frond are groups of sporangia called sori (Figure 25.23a). Most ferns are homosporous. Spores are produced by meiosis and are released into the air from the sporangium. Those that land on a suitable substrate germinate and form a heart-shaped gametophyte, or prothallus, which is attached to the ground by thin filamentous rhizoids (Figure 25.23b). Gametophytes produce both antheridia and archegonia. Like the sperm cells of other pterophytes, fern sperm have multiple flagella and must swim to the archegonium, which releases a chemoattractant to guide them. The zygote develops into a fern sporophyte, which emerges from the archegonium of the gametophyte. Maturation of antheridia and archegonia at different times encourages cross-fertilization. The full life cycle of a fern is depicted in Figure 25.24.

The Importance of Seedless Plants
Mosses and liverworts are often the first macroscopic organisms to colonize an area. Once mosses and liverworts are established, they provide food and shelter for other plant and herbivorous animals. In turn, predators feed on the herbivores.
Ferns contribute to the environment by promoting the weathering of rock, accelerating the formation of topsoil, and slowing down erosion of the soil. The water ferns of the genus Azolla harbor nitrogen-fixing cyanobacteria and restore this important nutrient to aquatic habitats.
Seedless plants have historically played a role in human life with uses as tools, fuel, and medicine. For example, dried peat moss, Sphagnum, is commonly used as fuel in some parts of Europe, and are often cultivated with cranberry and blueberry bushes. In addition, the ability of Sphagnum to hold moisture makes the moss a common soil conditioner. Even florists use blocks of Sphagnum to maintain moisture for floral arrangements!
By far the greatest impact of seedless vascular plants on human life, however, comes from their extinct progenitors. The tall club mosses, horsetails, and tree-like ferns that flourished in the swampy forests of the Carboniferous period gave rise to large deposits of coal throughout the world. Coal provided an abundant source of energy during the Industrial Revolution, which had tremendous consequences on human societies, including rapid technological progress and growth of large cities, as well as the degradation of the environment. Coal is still a prime source of energy and also a major contributor to global warming.
