
4
Classifying Living & Fossil Hominids
Learning Objectives:
- Outline primate characteristics and basis of classification
- Describe living hominid behavior and social organization
- Explain principle anatomical trends in hominin evolution and within genus Homo
- Summarize fossil evidence for early hominin evolution
- Discuss theories of relationships and models of emergence for hominin ancestors in the human lineage
The Living Primates
In order to understand the exact place of humans among the animals, it is helpful to understand how we are taxonomically classified. In the eighteenth century, Carl von Linné placed humans together with other similar-looking organisms in the taxonomic order Primates. Modern molecular biology has reinforced our place with primates, as humans, monkeys and apes share the vast majority of their DNA, with chimpanzees sharing between 97 to 99% genetic identity with humans.
Primatology is the scientific study of behavior, biology, evolution, and taxonomy of primates. The focus is on studying the common links between humans and nonhuman primates. It is believed that by understanding our closest animal relatives, we might better understand the characteristics and deep history that we share. Through the study of primates we can learn what it means to be human, as opposed to being a primate, and further, as opposed to other mammals or animals in general. We can also learn what traits are shared between all of the primates, what traits some primates have lost, and new traits that primates developed in order to survive in different environments. We can also learn about our past, how we’ve evolved, who we’ve become, and what forces have shaped us into the highly unique animal we’ve become today.
Primatologists commonly use three methodological approaches in their research. Field study (Figure 4.1) is done in natural environments, in which scientific observers watch primates in their natural habitat. Laboratory study is done in controlled lab settings
Figure 4.1: Primatologist Claudia Wilke studying communication and cooperation among chimpanzees in the Kibale Forest, Uganda
where scientists are able to perform controlled experiments that focus on the learning capabilities and behavioral patterns of the animals. In semi-free ranging studies, scientists are able to watch how primates might act in the wild but have easier access to them, and the ability to control their environments.
Primate characteristics
Defining the term primate can be challenging at times because this biological order of animals is not defined by one fixed set of characteristics. However, there are some general features that most have in common.
Many primates are arboreal, meaning that they live in trees. Because of their arboreal adaptations, primates typically have the ability to grasp with their hands and sometimes the feet and tail as well. Grasping in this manner is known as prehensile capability (Figure 4.2). Primates typically have opposable toes and thumbs (Figure 4.3), which provide a firm, tight grasp on tree branches.
Figure 4.2: Red howler monkey with prehensile capabilities in hands, feet and tail
Figure 4.3: Opposability in the fingers and toes of an orangutan
Primates share a generalized skeletal structure that allows for greater flexibility in their joints, limbs and torso (Figure 4.4). Bones in the hands and feet are separate, increasing the range of motion, as well as the power and precision when manipulating objects. The ends of primate fingers and toes have dermal ridges that are very sensitive to textures of objects, such as tree branches and potential foods. They usually have flat fingernails (Figure 4.5) rather than curved, pointy claws. Nails help primates manipulate objects more easily and protect the sensitive ends of fingers and toes.
Figure 4.4: Generalized primate skeletal structure of an orangutan

Figure 4.5: Nails on chimpanzee hand and foot
The primates’ adaptation to their way of life in the trees coincided with changes in the form and function of their senses. Primates have forward-facing eyes on the front of the skull, producing binocular, or overlapping, fields of vision (Figure 4.6), allowing them to be very adept at judging depth, direction, distance and the relationship of objects in space. In addition, their vision is very colorful and sharp. With their dependence on enhanced vision, they have a reduced sense of smell and hearing, which is evident in their smaller and less projecting snout when compared with other mammals. Primate dentition reflects their dietary diversity with different tooth types to slice, puncture, crush and grind the wide range of foods being eaten.
Figure 4.6: Primate binocular vision produces a larger overlapping field of vision than other animals, which increases depth and spatial perception of objects.
Primates tend to give birth to smaller numbers of offspring, most often one at a time, with birthing spaced out over longer periods (Figure 4.7). The offspring take significant time to develop and have a longer lifespan than many other animals. This is in part due to a larger and more complex brain relative to their body size when compared with the rest of the animal kingdom (Figure 4.8). A larger and more complex brain not only provided primates with a greater degree of efficiency in the daily struggle for survival but also gave them the basis for more sophisticated thought process.
Figure 4.7: Adult female olive baboon with juvenile
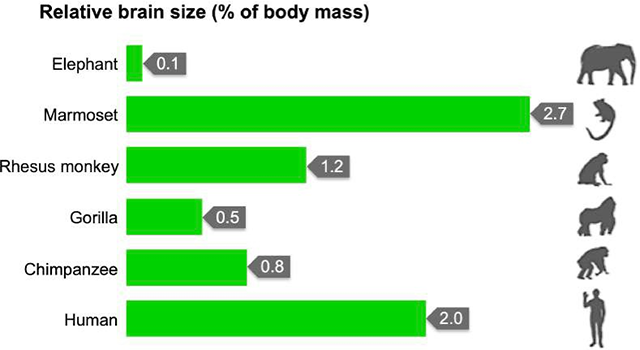
Figure 4.8: Relative comparison of primate brain size to body mass
Primate classification
Similarities between humans and primates are evident in not only the shared physical features, but also in the placement and degree of relatedness of humans and primates in Linnaean taxonomy (Figure 4.9).
Figure 4.9: Linnaean taxonomy of major family groups of living primates
Figure 4.10: Distribution of living primates in a wide range of environments
Living primate diversity
Today, there are over 200 species of known primates living in various parts of the world (Figure 4.10). These species are divided into two taxonomic groups: Strepsirrhini and Haplorrhini. Strepsirrhines include arboreal species of lemurs, pottos, galagos and lorises (Figure 4.11) found in tropical Africa, Madagascar, India and Southeast Asia. Strepsirrhines share many ancestral characteristics, but at the same time, some extreme specializations for particular modes of life. They rely heavily on a more pronounced sense of smell, an adaptation reflected in their enlarged nasal passages and a rhinarium, a moist, fleshy pad at the end of their nose similar to a cat or dog. Strepsirrhines use a specialized set of teeth, known as a tooth comb, in the front, lower part of the mouth that angle forward, and a toilet claw on the second toe on the hind foot in grooming. Their eyes contain a reflective layer to improve their night vision for nocturnal activity, and their eye sockets include a ring of bone around the eye rather than being fully enclosed. They also produce their own vitamin C, rather than obtaining it solely from their diet of fruits, leaves and insects.
Haplorhines are more widely distributed geographically and include many more species. These species are further divided into three taxonomic groups: Tarsiidae, Platyrrhini and Catarrhini. Tarsiers (Figure 4.12) are only found in the island areas of Southeast Asia and are small animals with enormous eyes, in some cases larger than its entire brain, to help them see well at night. They can rotate their heads around 180 degrees, which is necessary because they cannot move their eyes side to side. They also have an excellent sense of hearing, which aids in avoiding predators and finding food. Tarsiers are strict carnivores, feeding on insects, birds, and lizards.
Figure 4.11: Strepsirrhines include species such as the (a) ring-tailed lemur ( Lemur catta ), (b) red slender loris ( Loris tardigradus ), (c) Senegal galago ( Galago senegalensis ) and (d) potto ( Periodicticus potto )
Figure 4.12: Tarsiidae include species such as the Philippine tarsier ( Carlito syrichta )
Figure 4.13: Platyrrhines, or New World monkeys, include species such as the (a) white-faced saki ( Pithecia pithecia ), (b) golden lion tamarin ( Leontopithecus rosalia ), (c) black howler monkey ( Alouatta caraya ), and (d) brown capuchin ( Sapajus apella)
Platyrrhines, or New World monkeys, are found in the tropical regions of Central and South America and Mexico. This group includes monkeys that are smaller to mid-sized, such as sakis, tamarins, capuchins, and howlers (Figure 4.13). Most are quadrupedal, moving using all four feet, and primarily arboreal and diurnal, or active during the day. They are the only monkeys to have prehensile tails. Their noses are flatter than those of other primates, with sideways-facing nostrils. Platyrrhines are omnivores, meaning they eat plants and animals. They have an extra set of upper and lower molars that are effective in processing their foods, which primarily consist of fruits, nuts, insects, flowers, bird eggs, spiders, and even small mammals.
Catarrhines include three taxonomic families: Cercopithecidea, Hylobatidae and Hominidea. They display much more morphological and behavioral diversity than New World monkeys. Except for humans, they are the most widely distributed of all living primates. They are found throughout sub-Saharan Africa and Southern Asia, ranging from tropical jungle habitats to semiarid desert and even to seasonally snow-covered areas in northern Japan. Most are quadrupedal and primarily arboreal. All catarrhines have nostrils that face downwards and do not have prehensile tails.
Cercopithecids, or Old World monkeys, are found in Africa and Asia, inhabiting numerous environments that include tropical rain forests, savannas, shrublands, and mountainous terrain. This group includes monkeys that are medium to larger in size, such as macaques, colobus monkeys, baboons, and mandrills (Figure 4.14). Some cercopithecids, like the colobus monkey, are arboreal, but most, like baboons and many macaques, are terrestrial, meaning they primarily live on the ground.
Figure 4.14: Cercopithecids, or Old World monkeys, include species such as (a) gelada baboons ( Theropithecus gelada ), (b) Japanese macaques ( Macaca fuscata ), (c) mandrills ( Mandrillus sphinx ), and (d) eastern back-and-white colobus monkeys ( Colobus guereza )
All cercopithecids have tails; however, they are not prehensile. They also have very distinctive, and at times very colorful, rough, hardened skin pads on their buttocks, or ischial callosities, that enable the monkeys to sleep sitting upright on thin branches, beyond reach of predators, without falling. Most cercopithecids are at least partially omnivores, but all prefer plant matter, which forms the bulk of their diet. Most are highly opportunistic, primarily eating fruit, but also consuming almost any food items available, such as leaves and bulbs, insects, snails, and small mammals.
Hylobatids, commonly referred to as lesser apes, are found in the tropical forests of islands and mainland of Southeast Asia and include species of siamangs and gibbons (Figure 4.15). They are medium to larger in size, lack tails, and have ischial callosities. Their arms are longer than their legs, and they have elongated, thin hands and feet, all adaptations for brachiation (Figure 4.16), a suspensory hand-over-hand movement swinging from branch to branch. They are strongly territorial, and defend their
Figure 4.15: Hylobatids, or the lesser apes, include species such as (a) the siamang ( Symphalangus sydactylus ) and (b) the lar gibbon ( Hylobates lar )
Figure 4.16: Young Lar gibbon brachiating though the trees
boundaries with vigorous visual and vocal displays. Some species have an enlarged throat sac, which inflates and serves as a resonating chamber when the animals call to each other. This structure can become quite large in some species, sometimes equaling the size of the primate’s head. Their diets are about 60% fruit-based, but they also consume twigs, leaves, insects, flowers, and occasionally bird eggs.
Living Hominid family
Hominids, more commonly known as great apes, include orangutans, gorillas, chimpanzees, and humans. They are the largest in body size of all primates with notable differences in size between males and females although the degree of difference varies greatly among species. Hominids do not have tails, nor do they have ischial callosities. All hominids, except for humans, have arms that are longer than their legs. They use their longer arms for knuckle walking (Figure 4.17), where the arms and knuckles of the hands support the weight of the torso as they move around. Their large toe and thumb are opposable, also an exception in humans, and all fingers and toes have flattened nails. All hominids have larger brains to body mass ratios, which reflect the complexity behind their cognitive abilities.
Orangutans (Figure 4.18) include three living species currently only found in the rainforests of the Indonesian islands of Borneo and Sumatra. They are slow, cautious climbers whose method of movement can best be described as “four-handed,” a tendency
Figure 4.17: Adult male Western lowland gorilla knuckle walking
to use all four limbs for grasping and support. They are the most arboreal of the great apes and spend most of their time in trees. They are very large animals with males and females greatly differing in size and appearance. Males weigh over 90 kg (200 lb), while females are usually less than 45 kg (100 lb). Dominant adult males have distinctive cheek pads and throat pouches, used when producing long calls that attract females and intimidating rivals. Younger males do not have these characteristics and resemble adult females.
Orangutans are the most solitary of the great apes, with social bonds occurring primarily between mothers and their dependent offspring, who stay together for at least the first two years. Orangutans use a variety of sophisticated tools to obtain food resources and construct elaborate sleeping nests each night from branches and foliage. Fruit is the most important component of an orangutan’s diet; however, they will also eat vegetation, bark, honey, insects and even bird eggs.
Figure 4.18: An (a) adult male Bornean orangutan ( Pongo pygmaeus ) with cheek pads and throat pouch, and (b) and adult female with a juvenile
Gorillas (Figure 4.19) are the largest of all living primates and currently they are confined to forested areas of western and equatorial Africa. There are two species of gorilla: the Western species, which is further divided into Western Lowland and Cross River gorillas, and the Eastern species, which are also further divided into Eastern Lowland and Mountain gorillas. Lowland gorillas live in dense forests and lowland swamps and marshes. Mountain gorillas inhabit moist tropical and subtropical mountainous forests with low-level cloud cover, and Cross River gorillas live in low-lying and temperate forested hills and mountains. Gorillas live in groups called troops. Troops tend to be made of one adult male, multiple adult females and their offspring.
Gorillas exhibit extreme physical differences between males and females. Adult males are on average 1.6 m (5.2 ft) tall and weigh approximately 177 kg (390 lb), while adult females weight on average 80 kg (176 lb) with a height of approximately 1.4 m (4.6 ft). Because of their size, gorillas construct nests on the ground for daytime and night use. Gorillas move around by knuckle walking, although they sometimes walk upright for short distances while carrying food or in defensive situations. A gorilla’s day is divided between rest periods and travel or feeding periods. Diets differ between and within species. Mountain gorillas mostly eat foliage, such as leaves, stems, pith, and shoots, while fruit makes up a very small part of their diets. Lowland gorillas have more diverse diets, with leaves and piths commonly eaten, but fruits making up as much as 25% of their diets. The DNA of gorillas is highly similar to that of humans, from 95 to 99% depending on what is included, and they are the next closest living relatives to humans after the chimpanzees.
Figure 4.19: An (a) adult male Eastern Mountain gorilla ( Gorilla gorilla ) compared to (b) an adult male Western Lowland gorilla ( Gorilla berengei ), and (c) and adult female and infant Eastern Mountain gorilla compared to (d) an adult female and infant Western Lowland gorilla
The most well-known of all nonhuman primates are chimpanzees, found in the forests and savannas of equatorial Africa. There are two species of chimpanzee: the common chimpanzee and the bonobo. Common chimpanzees (Figure 4.20), in many ways, are structurally similar to gorillas, with corresponding limb proportions and upper body shape. Chimpanzee males tend to weigh around 50 kg (110 lb) and stand about 1.4 m (4.6 ft) in height, with the females being slightly smaller in body size. The physical strength of chimps is around 1.5 times greater than humans. Chimpanzees are adapted for both arboreal and terrestrial movement. The hands have long fingers with short thumbs and flat fingernails, and the feet are adapted for grasping with an opposable large toe. Arboreal locomotion consists of vertical climbing and brachiation. On the ground, chimps move both Chimpanzees live in communities that typically range from 20 to more than 150 members, but spend most of their time traveling in small, temporary groups consisting of a few individuals, which may consist of any combination of age and sex. quadrupedally by knuckle-walking and frequently upright for short distances.
Chimpanzees are highly adaptable and live in a variety of habitats, including rainforest, mountainous cloud forest, swamp forest and dry woodland-savanna landscapes. They are omnivores, but prefer fruit above all other food items. They also eat leaf buds and blossoms, seeds, stems, pith, bark, and resins. Nearly all chimpanzee populations have been recorded using tools, modifying sticks, rocks, grass and leaves and using them for acquiring honey, termites, ants, nuts and water. The species has also been found creating sharpened sticks to spear small mammals and have been known to hunt and consume lower-order primates.
Figure 4.20: An (a) adult male and (b) an adult female and infant chimpanzee
( Pan troglodytes )
Bonobos (Figure 4.21) are found in the forests of the Congo River Basin of Central Africa. Bonobos can be distinguished from common chimpanzees by their relatively long legs, pink lips, dark face and tail-tuft through adulthood, and parted long hair on their head. They also have a slim upper body, narrow shoulders, thin neck, and long legs when compared to the common chimpanzee. Although chimpanzees are typically larger in height and weight than bonobos, the two species tend to broadly overlap in body size. They are both terrestrial and arboreal, using the same modes of movement as chimpanzees.
Bonobos live in communities of approximately 100 members, which will split into small groups during the day while looking for food, and then come back together to sleep. They sleep in nests that they construct in trees. They are also omnivores, with fruit making up over 50% of their diet. They also eat leaves, honey, bird eggs, meat from small mammals and insects. Sexual activity generally plays a major role in bonobo society, used as a greeting, a means of forming social bonds, and as a means of conflict resolution and reconciliation.
Figure 4.21: An (a) adult male and (b) adult female bonobo ( Pan paniscus )
Our Fossil Ancestors
You may have noticed there is only one species of human being on earth: Homo sapiens. As we have seen above, humans are just one small branch on the primate family tree. In comparison, many other primates have more than one extant species within their grouping, for instance, two species of gorilla, 18 species of gibbons, and over 130 species of Old World monkeys. Ancestral humans used to live among other species of human. New species appeared at various points in history as ancestral humans adapted to their environment. The species were somewhat different from one another, but genetic evidence has shown that at times they may have interbred. The earliest of these species developed different traits compared with the other apes. Ultimately, they laid an evolutionary path that helped to define who we are today, as Homo sapiens.
Paleoanthropology is the study of the origins and development of early humans and their ancestors and focuses on the fossil record of humans and non-human primates. Paleoanthropologists need to be well versed in other scientific disciplines and methods, including ecology, biology, anatomy, genetics, and primatology. Paleoanthropologists have various interpretations of fossil classifications and lineages that have evolved over the millennia, but there is a general consensus that Homo sapiens last shared an ancestor with the great apes approximately 6.0 to 8.0 million years ago. These ancestral species are classified as hominids, from family Hominidae, which currently includes the genera:
- Pongo – the Bornean, Sumatran and Tapanuli orangutan
- Gorilla – the Eastern and Western gorilla
- Pan – the common chimpanzee and the bonobo
- Homo – modern humans and their extinct human-like ancestors.
Several revisions in primate classification have caused the use of the term hominid to vary over time (Figure 4.22). Its original meaning referred only to humans and their closest extinct relatives. But, the current meaning of hominid includes all of the great apes including humans. The term hominin, from the taxonomic tribe classification Hominini, is used when referring to modern humans and our close extinct relatives.
Evidence of hominin activity between 8.0 and 2.5 million years ago typically consists of fossil remains of bones and teeth. This is one reason that specialists from biological anthropology (e.g., osteologists, functional anatomists) are essential to understanding and explaining processes of evolutionary adaptation and variation.
Figure 4.22: Changes in classification and use of the terms “hominid” and “hominin”
Hominin evolutionary trends
Bipedalism, or the ability to habitually walk on two legs, began in human evolution as early as 7.0 million years ago. This form of locomotion has led to morphological changes (Figure 4.23) in the human skeleton including changes to the arrangement and size of the foot, pelvic size and shape, knee size and position, leg length, shape and orientation of the vertebral column, and positioning of the skull.
Human walking is functionally about 75% less costly than both quadrupedal and bipedal walking in chimpanzees. Some hypotheses have supported that bipedalism increased the energetic efficiency of travel and that this was an important factor in the origin of bipedal locomotion. While humans save more energy than quadrupeds when walking, this is not the case when running, as human running is functionally 75% less efficient than human walking.
Figure 4.23: Morphological changes in the human skeleton related to bipedalism
Human feet evolved enlarged heels to bear the body weight that also increased over time. The human foot evolved as a platform to support the entire weight of the body, rather than acting as a grasping structure, as it did in early hominids. As a result, modern humans have smaller toes than their bipedal ancestors. This includes a non-opposable first toe, which is relocated in line with the other toes. When non-human hominids walk upright, weight is transmitted from the heel, along the outside of the foot, and then through the middle toes, while a human foot transmits weight from the heel, along the outside of the foot, across the ball of the foot and finally through the first toe. This transference of weight contributes to energy conservation during walking.
Human hip joints are larger than in quadrupedal ancestral species to better support the greater amount of body weight passing through them, as well as having a shorter, broader shape. This change in shape brought the vertebral column closer to hip joints and increased surface area for ligaments supporting the abdominal viscera and gluteus muscles to attach, providing a stable base for support of the trunk while walking upright and while standing on one leg. The sacrum has also become broader, increasing the diameter of the birth canal and allow for rotational movement of the infant during birth.
Human knee joints are enlarged for the same reason as the hip joint – to better support an increased amount of body weight. The changing pattern of the knee joint angle in humans decreases energy lost by vertical movement of the center of gravity. Humans walk with their knees kept straight and the thighs bent inward so that the knees are almost directly under the body, rather than out to the side, as is the case in ancestral hominids. This type of gait also aids balance.
Increasing leg length in the evolution of bipedalism changed how leg muscles functioned in upright gait. In humans the “push” for walking comes from the leg muscles acting at the ankle. A longer leg allows the use of a natural swing of the limb so that, when walking, humans do not need to use muscles to swing the other leg forward for the next step.
The human vertebral column takes a forward bend in the lumbar region (lower torso) and a backward bend in the thoracic region (upper torso). Together the lumbar and thoracic curves bring the body’s center of gravity directly over the feet. In humans, the foramen magnum, an opening for passage of the spinal cord, is located at the base of the skull. This places most of the weight of the head behind the vertebral column, which is influences balance and posture during movement.
There is a general pattern of reduction in the size of the teeth, jaws and face. Back teeth, or molars, with thick enamel became highly specialized for grinding foods, while the front teeth, the incisors and canines, progressively became smaller and less specialized for slicing and cutting foods. As the size of teeth changed, the jaw also became smaller, lessening facial prognathism, or the protrusion of the upper and lower jaws.
Expansion of cranial capacity, or the size of the brain cavity in the skull, came later than bipedalism. Early hominins had brains almost equivalent with those of chimpanzees (320-480 cc), but fossil hominins with significantly increased cranial capacities (500-900 cc) did not appear until about 2.1 million years ago with the genus Homo. Expansion in cranial capacity developed along with a new pattern of cerebral blood circulation. Gravitational forces on blood draining from the brain differ in quadrupedal animals versus bipedal animals; therefore, when humans stand bipedally, most blood drains into veins at the back of the neck, which form a complex system around the spinal column. Savanna-dwelling hominins with this network of veins had a way to cool a bigger brain, allowing for an increased brain size, and further contributing to hominin flexibility in moving into and being active in new habitats with wide-ranging conditions.
Unlike other primates, hominins no longer used their hands in locomotion, bearing body weight or brachiating. The human hand is shorter than the foot, with straighter finger bones. Fossil hand bones about 2.5 million years ago reveal this shift in specialization of the hand from locomotion to fine manipulation and greater potential for tool use. Although no stone tools are recognizable in an archaeological context until 3.3 million years ago, we can infer that the tradition of making and using tools almost certainly goes back much earlier to a period of utilizing unmodified stones, organic perishable materials (e.g., wood) that would not be preserved in the fossil record, or animal teeth and horn that may not be easily recognized as tools.
Hominin Lineages
The hominin family tree is grouped by genus, or by related genera in the case of the earliest grouping. Exact relationships between these species are not well understood, and doubtless many more are waiting to be discovered. Currently, more than twenty different species of extinct hominins closely related to modern humans are recognized (Figure 4.24); however, some species merit special attention because they prompted paleoanthropologists to modify lineage relationships and our understanding of human evolution.
Earliest known Hominins
Molecular genetics tells us that humans, gorillas, bonobos and chimpanzees are all descended from a common ancestor that lived sometime between 7.5 million and 6.5 million years ago. All of the earliest known hominin fossils have been found in Central and Eastern Africa (Figure 4.25).
Sahelanthropus tchadensis lived sometime between 7.0 and 6.0 million years ago in Chad in West-Central Africa, making it the oldest known hominin to date. The species is represented by a mostly complete but distorted cranium and additional cranial fragments. Walking upright may have helped this species survive in diverse habitats, including forests and grasslands. Although there are no postcranial remains of
Figure 4.24: Timeline and evolutionary relationships of major hominin species
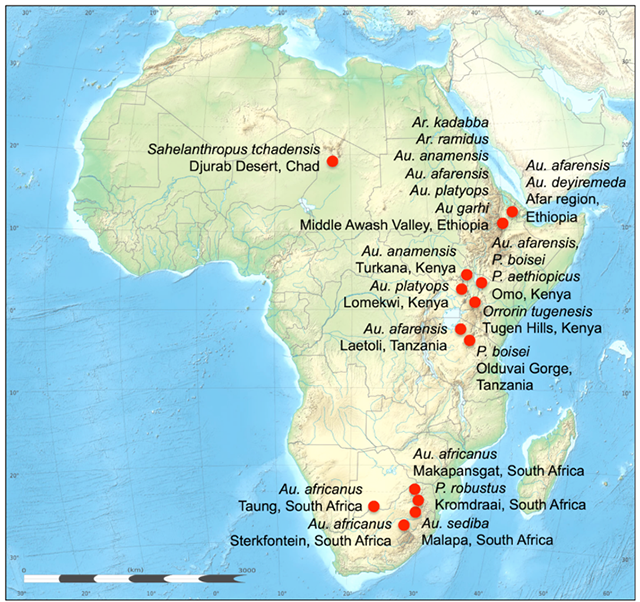
Figure 4.25: Sites of significant early hominin and australopithecine fossil discoveries
Sahelanthropus, we do know this species had a combination of ape-like and human-like features. Ape-like features included a brain slightly smaller than a chimpanzee’s, a sloping face, very prominent brow ridges, and elongated skull form. Human-like features included small canine teeth, a short mid-facial region, and the shape and central positioning of the foramen magnum under the skull suggest the ability to walk upright. Unfortunately, most of Sahelanthropus’ teeth are heavily worn; however, based on its environment and other early human species, it probably ate a mainly plant-based diet. This likely included leaves, fruit, seeds, roots, nuts, and insects.
Also living around 6.0 million years ago, Orrorin tugenensis is among the oldest early hominins. At least five individuals of this species were found near Lake Turkana in the Tugen Hills of Kenya, in Eastern Africa. Fossil remains of this species include bones of the arms and legs, parts of fingers and several teeth. Individuals of this species were approximately the size of a chimpanzee, standing less than 1 m (3.3 ft) tall, and had small teeth with thick enamel, similar to modern humans. From Orrorin ‘s low, rounded molars and small canine teeth, paleoanthropologists can infer that this species ate mainly a plant-based diet. The bones of the hand were curved like a living ape’s, suggesting that this species spent time in the trees; however, the most important fossil of this species is the upper portion of a femur, with the angled part more closely resembling that of a human. It formed a strong bridge with the hip to support the body’s weight, showing evidence typical of a biped – meaning Orrorin tugenensis individuals climbed trees but also probably walked bipedally.
Ardipithecus kadabba lived sometime between 5.8 and 5.5 million years ago in the Middle Awash Valley of Ethiopia in Eastern Africa. The species is represented by several teeth, partial arms, a clavicle, and hand and foot bones. Evidence for Ar. kadabba ’s bipedalism comes from a single toe bone that has a broad, robust appearance, suggesting its use in bipedal walking; however, the hand bones retain an ape-like curvature suggestive of time spent in trees. These individuals were probably similar in body and brain size to a modern chimpanzee and had larger canines. Instead of eating mostly fruit and soft leaves like chimpanzees, there is evidence that Ar. kadabba ate a variety of foods. The back teeth of Ar. kadabba are larger than a chimpanzee’s, but its front teeth are narrower suggesting this species did most of its chewing in the back of its mouth with a focus on hard-to-eat foods like fibrous nuts.
Over 100 specimens of Ardipithecus ramidus fossil remains have been discovered in Middle Awash Valley and Gona, Ethiopia in Eastern Africa. This species had an elongated skull, a sloping face, very prominent brow ridges, and small brain, slightly smaller than a chimpanzee’s and roughly 20% the size of the modern Homo sapiens brain. A good sample of canine teeth of this species indicates very little difference in size between males and females in this species. Ar. ramidus individuals were most likely omnivores, which means they enjoyed more generalized diet of plants, meat, and fruit, and did not seem to eat hard, abrasive foods like nuts and tubers, which is consistent with the dentition having thinner enamel than what is seen in earlier hominin species.
| Key Concepts: Earliest Known Hominins | |
|---|---|
|
Sahelanthropus tchadensis ‘Toumaï’ fossil |
Location: Djurab Desert, Chad
Date(s): 7.0-6.0 million years ago Habitat: Flooded grasslands, open woodlands Key traits:
|
|
Orrorin tugenensis ‘Original Man’ fossil
|
Location: Tugen Hills, Kenya
Date(s): 6.2-5.6 million years ago Habitat: Forest and wet woodlands Key traits:
|
|
Ardipithecus kadabba ‘Oldest Ancestor’ fossil |
Location: Middle Awash Valley, Ethiopia
Date(s): 5.8-5.5 million years ago Habitat: Woodlands, grasslands and open lake areas Key traits:
|
|
Ardipithecus ramidus ‘Ardi’ fossil |
Location: Middle Awash Valley and Gona, Ethiopia
Date(s): 4.5-4.3 million years ago Habitat: Woodlands, grasslands and open lake areas Key traits:
|
The ‘Ardi’ skeleton is the most complete early hominin specimen and reflects a particular mosaic of ape- and human-like characteristics not seen in later hominins. Ardi weighed about 50 kg (110 lb) and was up to 1.2 m (3.9 ft) tall. It is still a point of debate whether Ardi was consistently using bipedal movement, as she had both opposable big toes and thumbs suited for climbing trees. Curved bones in her toes and divergent larger toe are not characteristic of a biped, and her pelvis shows evidence of muscles used in climbing, and therefore was a quadruped when moving around in trees. However, remains of her legs, feet, pelvis, and hands also suggest that she walked upright when on the ground.
The Australopithecines
Australopithecus , which literally means ‘southern ape,’ is another hominin genus that evolved earlier than Homo. From paleontological and archaeological evidence, the Australopithecus species, referred to as gracile australopithecines, apparently evolved in eastern Africa around 4.0 million years ago before spreading throughout the continent and eventually becoming extinct around 1.2 million years ago. Australopithecus species played a significant role in human evolution, as the genus Homo may have descended from an australopithecine lineage around 2.5 million years ago.
Fossil remains indicate that these individuals generally had features that were more human-like than ape-like. They were fully bipedal, but did not walk identically to humans. Their height was shorter than the average modern human, standing 1.0 to 1.5 m (3.3 ft to 4.9 ft) tall, and they weighed between 30 and 55 kg (66 and 121 lb). They tended to have longer arms and greater sexual dimorphism, or distinct difference in size or appearance between the sexes, when compared to other hominins. Their brains were roughly 35% of the size of a modern human brain and more in the range of the brains of modern gorillas. Their facial features, however, looked more ape-like than human, with sloping faces and large, jutting jaws. Their molars were relatively large and had more enamel when compared with modern apes and humans, but their canines were relatively small with very little difference between males and females.
Currently, there are seven generally accepted species, Au. anamensis, Au. afarensis,
Au. platyops, Au. deyiremeda, Au. africanus, Au. garhi, and Au. sediba, belonging to the genus, all of which were found in Eastern and Southern Africa.
Australopithecus anamensis, living around 4.2 to 3.9 million years ago, is the oldest known Australopithecine. Over 20 individuals of this species are represented by almost 100 fragments of skull, teeth, arms and legs, and hands and feet found at Lake Turkana, Kenya and Middle Awash Valley, Ethiopia in Eastern Africa. Au. anamensis appears to have been habitually bipedal, although it retained long forearms and curved bones in the hand, indicating that individuals probably climbed trees as well. At about 1m (3.3 ft) in height, their body was approximately the size of a chimpanzee.
Key traits in the jaws and teeth indicate Au. anamensis was possibly the direct ancestor of Au. afarensis, and possibly the direct descendent of a species of Ardipithecus. Individuals had thickly-built, long, narrow jaws with their side rows of teeth arranged in parallel lines. They had very large canines and large, broad molars with very thick enamel. Their strong jaws combined with heavily enameled teeth suggest these individuals may at times have eaten hard, abrasive foods, but they likely were plant-eaters in general, relying on fruits and tough foods such as nuts and seeds.
Australopithecus afarensis is one of the longest-lived and best-known early hominin species, living around 3.9 to 2.8 million years ago. This species survived for more than 900,000 years, which is over four times as long as our own species has been around. Fossil remains were found primarily at Hadar in the Afar region of Ethiopia in Eastern Africa, but evidence has also been recovered from Koobi Fora and Lothagam, Kenya and Laetoli, Tanzania. This species is known from remains of more than 300 individuals, including adults and children. There is significant sexual dimorphism among these individuals with an average male height of 1.5 m (4.9 ft) and body mass of 45 kg (99 lb), while females were approximately 1.0 m (3.3 ft) with a body mass of 29 kg (64 lb).
The skeletal anatomy of the hands, feet, and shoulder joints indicate that Au. afarensis was to some extent efficiently grasping branches and climbing trees. The curvature of the finger and toes bones and the shape of the bones in the shoulder retain ape-like qualities. However, the loss of the ability to fully grasp with the foot using divergent large toe also suggests that Au. afarensis was no longer adapted to climbing in the same manner as earlier hominin species. There are a number of traits in the pelvis, legs and feet that strongly reflect almost exclusive bipedality. The blades of the pelvis are short and wide for stability during walking or running, and the femur angles in toward the knee from the hip. This last trait also allowed the foot to fall closer to the midline of the body, a very strong indication of habitual bipedalism, in much the same manner as modern humans. At Laetoli, thousands of footprints of various animals were preserved in an ancient layer of mud covered with volcanic ash. These included a trail of the earliest footprints clearly made by a group of hominins approximately 3.6 million years ago (Figure 4.26).
Au. afarensis adaptations for living both in the trees and on the ground helped them survive for almost a million years as climate and environments changed.
There are several traits that suggest Au. afarensis may be a direct descendant of Au. anamensis and may be ancestral to later species of Paranthropus, Australopithecus, and Homo. The individuals had apelike face proportions with a flat nose and a strongly projecting lower jaw. Their skulls indicate they had fairly small brains, about a third of the size of a modern human brain. Au. afarensis children grew rapidly after birth, similar to chimpanzees, and reached adulthood earlier than modern humans. This meant
Au. afarensis had a shorter period of growing up than modern humans have today, leaving them less time for parental guidance and socialization during childhood. The wear on their teeth indicate they ate soft, sugar-rich fruits, but their tooth size and shape suggest
Figure 4.26: Footprints of Au. afarensis 3.6 million years ago (top) along with an analysis of step (center) and stride lengths (bottom).
they could have also eaten hard, brittle foods too, possibly as ‘fallback’ foods during seasons when fruits were not available.
Australopithecus platyops, also known as Kenyanthropus platyops, living around 3.5 to 3.2 million years ago in Eastern Africa. Fossil remains of this species include a relatively complete cranium and teeth found in the LakeTurkana region in Kenya. This species inhabited the region at the same time as Au. afarensis. While Au. platyops lived at the same time as Au. afarensis, the molars of Au. platyops were smaller, indicating that the two species may have had different diets and therefore likely did not compete for the same types of food, though they were both probably largely plant eaters. Many scientists think Au. afarensis is the ancestor of the genus Homo species and therefore modern humans, but some scientists now feel the flat face and less-pronounced brow ridges of
Au. platyops seem more closely related to Homo.
Also living in Eastern Africa in the Afar region around 3.5 to 3.3 million years ago is Australopithecus deyiremeda . This recently discovered species is represented by a partial upper jaw, two lower jaws, and some cranial fragments. These fossil remains have some of the same characteristics as Au. afarensis, but there are also some differences such as a larger lower jaw and smaller teeth with thicker enamel, suggesting that this species had a diet made up of harder, tougher, and more abrasive foods than Au. afarensis.
| Key Concepts: The Gracile Australopithecines | |
|---|---|
|
Australopithecus anamensis ‘Lake’ fossil |
Locations: Middle Awash Valley, Ethiopia; Lake Turkana, Kenya
Date(s): 4.2-3.9 million years ago Habitat: Forest and woodlands around lakes Key traits:
|

Australopithecus afarensis ‘Lucy’ fossil
|
Location: Afar Region in Ethiopia
Date(s): 3.8-2.9 million years ago Habitat: Woodlands and open grasslands Key traits:
|
|
Australopithecus platyops ‘Flat-face’ fossil |
Location: Lake Turkana region, Kenya
Date(s): 3.5-3.2 million years ago Habitat: Woodlands Key traits:
|
|
Australopithecus deyiremeda ‘Close relative’ fossil |
Location: Lake Turkana region, Kenya
Date(s): 3.5-3.2 million years ago Habitat: Woodlands Key traits:
|
| Key Concepts: The Gracile Australopithecines | |
|
Australopithecus africanus ‘Mrs. Ples’ fossil |
Location: Taung, Sterkfontein and Makapansgat, South Africa
Date(s): 3.3-2.0 million years ago Habitat: Woodlands and open grasslands Key traits:
|
|
Australopithecus garhi ‘Surprise’ fossil |
Location: Bouri, Ethiopia
Date(s): 2.5 million years ago Habitat: Woodlands and open grasslands Tool culture: Early Mode I (Oldowan) Key traits:
|
|
Australopithecus sediba ‘Natural spring’ fossil |
Location: Malapa, South Africa
Date(s): 1.8 to 2.0 million years ago Habitat: Woodlands and open grasslands Key traits:
|
Australopithecus africanus lived sometime between 3.3 and 2.0 million years ago. Fossil remains, including the partial skeletons of two adults and multiple crania of adults and children, have been found at the sites of Taung, Sterkfontein, Makapansgat and Gladysvale in central South Africa. Like Au. afarensis of East Africa, this species represents viable candidates as ancestors of the genus Homo. Au. africanus was anatomically similar to Au. afarensis, with a combination of human-like and ape-like features. Compared to
Au. afarensis, Au. africanus had a rounder cranium with a higher forehead, less
pronounced brow ridge, slightly larger brain, smaller canines and large molars. Dental wear indicates these individuals ate tough foods but also had a very variable diet including softer fruits and plants. Au. africanus ape-like features include relatively long arms and a strongly sloping face that juts out from underneath the cranium with a pronounced jaw. The femur, foot and pelvis of Au. africanus indicate bipedalism, but shoulder and hand bones indicate they were also adapted for climbing in the same manner as Au. afarensis.
Australopithecus garhi dates to around 2.5 million years ago, and lived in the Middle Awash Valley, Ethiopia in Eastern Africa. The species is represented by one fossil cranium and four other skull fragments, although a partial skeleton found nearby, from about the same layer, is usually included as part of this species sample. The partial skeleton includes fragments of a femur that appears to be longer than those of other australopithecines; however, there is also evidence from this skeleton that indicates
Au. garhi still had long, powerful arms, a small cranial capacity, and strong facial prognathism. This suggests that the remains may represent a transitional stage between Australopithecus and Homo. Key traits in the teeth show that Au. garhi had the largest molars among the Australopithecines. These teeth also had thick enamel, indicating these individuals most likely ate tough foods similar to the diet seen in Au. africanus. However, A. garhi exhibits novel traits only otherwise seen in Paranthropus, such as a small sagittal crest, a ridge of bone running lengthwise along the midline of the top of the skull, indicating the presence of exceptionally strong jaw muscles.
Primitive-shaped stone tools were discovered with the Au. garhi fossils, dating back roughly 2.5 million years ago, along with animal bones that were cut and broken open with stone tools. It is possible, then, that this species was among the earliest to make the transition to stone tool technology and eating meat and bone marrow from large animals.
Australopithecus sediba lived in South Africa around 1.8 to 2.0 million years ago. The species is represented by six skeletons, which include an adult male, adult female, three infants, and a younger male child Au. sediba may have lived in savannas but ate fruit and other foods from the forests, a behavior similar to modern-day savanna chimpanzees.
Au. sediba is small in size, with long arms and legs, a narrow upper chest, and small cranial capacity, similar to other australopithecines, while details in the teeth and a broad lower chest resemble genus Homo. The pelvis and legs suggest that Au. sediba walked upright on a regular basis and that changes in the pelvis occurred before other changes in the body that are found in later specimens of Homo. However, the legs and feet point to a previously unknown way of walking upright – Au. sediba turned its foot inward with its weight focused on the outer edge of the foot.
Similarities and differences found in Au. sediba and Au. africanus suggests that Au. sediba was descended from Au. africanus. Alternately, Au. sediba may be closely related to another, still unknown species ancestral to genus Homo. Traits shared between Au. sediba and genus Homo indicate a closer relationship than any of the other australopithecines.
Paranthropus is a genus classification commonly used when referring to fossils also known as robust australopithecines. They are characterized by robust cranial and dental traits, such as sagittal crests, suggesting the presence of strong chewing muscles, and broad, grinding herbivorous molar teeth, which indicate a diet of tough vegetation, rather than hard foods, like seeds and nuts. They also share a wide, dish-shaped face with flaring cheekbones and a heavy brow ridge. The identification of robust forms of australopithecines emphasized an increase in diversity and complexity among hominins that led many paleoanthropologists to question previous ideas of a straightforward unidirectional evolution from an early ancestor to modern humans. These individuals lived between 2.7 and 1.1 million years ago in East and South Africa, when species of the genus Homo were prevalent, and probably descended from Au. afarensis and Au. africanus species. There are three generally accepted Paranthropus species, P. aethiopicus, P. boisei, and P. robustus, belonging to the genus.
Paranthropus aethiopicus lived in East Africa around 2.7 to 2.3 million years ago. The species is represented by two mostly complete skulls and a few jaw fragments.
P. aethiopicus has a strongly prognathic face, large molar teeth, a powerful jaw, and a well-developed sagittal crest, indicating huge chewing muscles, with a strong emphasis on the muscles that connected toward the back of the crest and created strong chewing forces on the front teeth. Many features of the skull are quite similar to Au. afarensis, and P. aethiopicus may be a descendent of this species. It is most likely the ancestor of
P. boisei, a r obust australopithecine species found later in Eastern Africa.
Paranthropus boisei lived between 2.3 and 1.2 million years ago in Eastern Africa. Adult males were larger on average than females, with males weighing about 49 kg (108 lb) and standing about 1.37 m (4.5 ft) tall, while females weighed about 34 kg (75 lb) and were 1.24 m (4.0 ft) tall. Like other Paranthropus species, P. boisei had the same facial and dental characteristics and jaw for chewing. However, this species had even larger molar teeth (Figure 4.27) than P. robustus, a much flatter, larger-brained skull than
Figure 4.27: Paranthropus boisei mandible and dentition (left) compared to Homo sapiens mandible and dentition (right).
P. aethiopicus, and the thickest dental enamel of any known early hominin. Along with fruits and nuts, their diet was largely based on grasses, sedges and tubers, making this species the largest consumer of plant matter than any other hominin studied to date.
Key Concepts: Robust Australopithecines

Paranthropus aethiopicus
‘Black Skull’ fossil
Date(s): 2.7-2.3 million years ago
Habitat: Open grasslands
Key traits:
- Heavy robust skull with sagittal crest and powerful jaw
- Wide, dish-shaped face with flaring cheekbones
- Small brain (~410 cc)
- Massive molar teeth
Paranthropus boisei
‘Nutcracker’ fossil
Date(s): 2.3-1.2 million years ago
Habitat: Open grasslands
Key traits:
- Heavy robust skull with sagittal crest and powerful jaw
- Wide, dish-shaped face with flaring cheekbones
- Small brain (~510 cc)
- Massive molar teeth; thickest enamel of all known hominins
 Paranthropus robustus
Paranthropus robustus‘Beside Man’ fossil
Date(s): 2.0-1.4 million years ago
Habitat: Open grassland and
Key traits:
- Heavy robust skull with sagittal crest and powerful jaw
- Wide, dish-shaped face with flaring cheekbones
- Small brain (~530 cc)
- Massive molar teeth
- Notable size difference between males and females
Paranthropus robustus lived in South Africa around 2.0 to 1.4 million years ago. P. robustus males may have stood only 1.2 m (4.0 ft) tall and weighed 54 kg (120 lb), while females stood just under 1 m (3.2 ft) tall and weighed only 40 kg (90 lb), indicating a large difference in size and weight between the sexes. P. robustus also had very large molar teeth with thick enamel and focused their chewing in the back of the jaw. Large flaring cheek bones allowed the passage of large chewing muscles to the jaw and gave P. robustus individuals their characteristically wide, dish-shaped face. These features allowed individuals to crush and grind hard foods such as nuts, seeds, roots, and tubers; however, P. robustus didn’t just eat tough foods. This species may have been more of a dietary generalist, also eating variety of other foods such as soft fruits and possibly young leaves, insects, and meat. While no stone tools are associated with P. robustus fossils, bone fragments show that these individuals probably used bones as tools to dig in termite mounds. Through repeated use, ends of these tools became rounded and polished. Termites are rich in protein and would have been a nutritious source of food.
Early Genus Homo
About 1.8 to 1.6 million years ago, the climate over most of Africa and Asia became drier and more seasonal with extensive open grasslands. In Asia, seasonal changes included three cold glacial periods with harsh, winter temperatures. The cooling and drying that occurred in these glacial periods brought an expansion of open habitats, with grasslands and mixed steppes. Early genus Homo was the first group to take advantage of these more arid and open environments. In the dry savannahs, plant tubers would probably have been an important part of the diet. These tough vegetables may have been processed using their improved tool technology as their smaller molar teeth imply that they ate foods that required less chewing. Their narrower pelvis and rib cage suggests that they had a smaller gut than earlier species such as A. afarensis. The development of a smaller gut and a bigger brain required more nourishing food, and this suggests that they may have included more meat in their diets.
Fire may have been used as long as 1.5 million years ago for cooking and warmth but whether this was a controlled use of fire is not certain. Charcoal, burnt earth, and charred bones found associated with early genus Homo fossils in Africa may have resulted from naturally occurring fires rather than from intentionally lit and controlled fires. However, discoveries in Wonderwerk Cave, South Africa, suggest controlled use of fire may have been occurring by 1.7 million years ago. Stratified deposits contain burnt stones and bones and layered traces of ash that indicate repeated burning events. This suggests that individuals regularly gathered around the fire to prepare and cook food and possibly also for social reasons.
The number of species within the genus Homo remains under debate, as classifying the genus Homo into species and subspecies is subject to incomplete information and is often highly problematic. Currently, there are over 15 different taxonomic species names used. Some of which are closely related to each other and have been considered to be subspecies of H. sapiens. This has led to using common names, such as “Neanderthal” to avoid trinomial taxonomic species names or the ambiguity of classifying groups with uncertain placement. Some extinct species in the genus Homo are only recently discovered (e.g., Denisova hominins) and do not as yet have consensus binomial taxonomic species names. There has historically been a trend to suggest the existence of new genus Homo species based on as little as individual fossil teeth. However, a minimalist approach to human taxonomy recognizes two early genus Homo species:
H. habilis and H. erectus.
Homo habilis lived predominantly in East Africa (Figure 4.28) between 2.4 and 1.4 million years ago. The brain of H. habilis was on average 50% larger than what is seen in australopithecines. This is reflected in significant changes to the shape of the skull, which is more rounded. There is less prognathism, and the brow ridges are smaller and arched. However, limb proportions are similar to those of the earlier australopithecine ancestors, with shorter legs and longer arms. H. habilis was smaller than modern humans, on
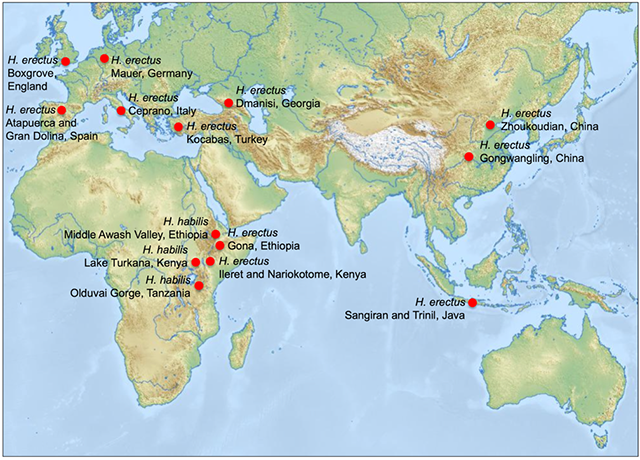
Figure 4.28: Sites of significant early genus Homo fossil discoveries
average standing no more than 1.3 m (4.3 ft). They still retained slight curvature in bones of the hand, but the fingers clearly show full capability of precision grip, where the tips of the fingers and thumb come together for fine manipulation of objects (Figure 4.29).
H. habilis lived in a predominantly grassland environment. The climate was becoming cooler and drier and this may have been the impetus for new feeding strategies that included scavenging and tool use. Dental microwear studies suggest that the diet of
H. habili s was flexible and versatile and that they were capable of eating a broad range of foods, including some tougher foods like leaves, woody plants, and some animal tissues, but that they did not routinely consume or specialize in eating hard foods like brittle nuts or seeds, dried meat, or very hard tubers. Another line of evidence for the diet of
H. habilis comes from some of the earliest cut- and percussion-marked bones, dating back to 2.6 million years ago. These traces of tool use on bones of larger animals are considered direct evidence of meat and marrow eating.
While paleoanthropologists used to place H. habilis as the direct ancestor to H. erectus, a relatively late 1.4 million-year-old H. habilis and a 1.6 million-year-old H. erectus from the same area of northern Kenya challenged the conventional view that these species evolved one after the other. Instead, this evidence, along with other H. habilis and H. erectus fossils that overlap in dating, demonstrate that they co-existed in Eastern Africa for more than half a million years.
There are some researchers who split the taxonomic classification of H. habilis, classifying some fossils as a separate species, Homo rudolfensis, based on the shape and size of the cranium and molar teeth. It remains an open question whether the fossil evidence is sufficient for suggesting a separate species, and if so whether this species should be classified within the genus Homo or genus Australopithecus, and if as Homo, whether it should be included under H. habilis, or even a more morphologically diverse species, Homo erectus.
Figure 4.29: Example of power grip (left) and precision grip (right).
| Key Concepts: Early Genus Homo | |
|---|---|
|
Homo habilis ‘Handy Man’ fossil |
Location: East Africa
Date(s): 2.4-1.4 million years ago Habitat: Wet grasslands, open woodlands and savannas Tool industry: Mode 1 (Oldowan) Key traits:
|
|
Homo erectus ‘Upright Man’ fossil |
Location: Africa, Asia, Southeast Asia, Europe
Date(s): 1.9 million – 300,000 years ago Habitat: Tropical forests and grasslands near streams and lakes Tool industry: Mode 2 (Acheulean) Key traits:
|
The core group within the species Homo erectus lived 1.9 to 1.5 million years ago, although some taxonomic classifications include additional individuals, extending their range up to about 300,000 years ago. H. erectus may have been the first hominin to migrate out of Africa. Fossil remains for H. erectus have been discovered at sites such as Gona in Ethiopia, Gran Dolina in Spain, Dmanisi in the Republic of Georgia, Zhoukoudian in China, and Sangiran in Java, Indonesia (Figure 4.28). Some researchers classify African H. erectus as Homo ergaster and consider this species to be the common ancestor of two groups: Eurasian H. erectus and H. sapiens. However, a skull discovered in 2000 at the site of Ileret, Kenya blurs the distinction between Eurasian H. erectus and African H. ergaster. It shows features that had previously only been found in Eurasian H. erectus specimens. This mix of traits caused some experts to rethink the whether these should be recognized as separate species.
African H. erectus was tall with a slender, long-legged body, which may have been an adaptation to maximize cooling of the body in a hot, dry environment. In addition, the body may have been relatively hairless as a way of improving body cooling by sweating. Unlike earlier species, the legs were much longer than the arms, so the limb proportions were similar to those of modern humans. All tree climbing adaptations of earlier species had been lost and had given way to a long-legged striding walk that was an efficient way to move about and made it easier to travel longer distances. Changes to the shoulders, chest and waist enabled the body to stay balanced during prolonged running. Fossil footprints made by H. erectus at Ileret, Kenya (Figure 4.30), dated to 1.5 million years ago, are the oldest known evidence of an essentially modern human-like foot anatomy and differ from the Laetoli footprints left by australopithecines 3.6 million years ago. This also makes them the oldest surviving footprints made by a human species.
The skull of African H. erectus had developed into a more human-like shape including a higher, more rounded braincase. Unlike modern humans, the cranium had a moderate post-orbital constriction, or the narrowing of the cranium just behind the eye orbits. The face projected outward but to a smaller degree than in earlier ancestors. A distinct double-arched brow ridge lay above the eyes, and a relatively distinct groove was located between the brow ridge and forehead. For the first time, the nose was human-like, now projecting outward rather than having a flattened appearance. The lower jaw was shorter and more lightly built than those of earlier species, resulting in a shorter face. The side rows of teeth were much further apart at the back of the jaw than at the front, forming a V-shaped pattern. The premolar and molar teeth were smaller and more human-like than those of earlier species., while the canine teeth were short and blunt, like those of modern

Figure 4.30: Footprints of Homo erectus, 1.5 million years ago at Ileret, Kenya, indicating modern human-like foot anatomy
humans. Vertebrae from the neck region of the spinal column indicate that the spinal cord was thinner than that of modern humans. This may indicate that African H. erectus had limited speech abilities due to the lack of nerves needed for complex control of breathing while speaking.
The most complete example of African H. erectus is known as ‘Nariokotome Boy’, who lived about 1.5 million years ago. He was about 8 to 10 years of age when he died but was already 1.6 m (5.2 ft) tall and may have reached 1.85 m (6.0 ft) as an adult. Almost 90% of his skeleton was recovered and has provided valuable information on this species’ body size, proportions, growth and development. The ‘Nariokotome Boy’ had a tall, slender body adapted for striding out across the extensive savannah plains. He also had a more human-like face with a nose that projected outwards and a larger braincase.
A female African H. erectus pelvis from Gona, Ethiopia, dated to 1.8 million years ago, suggests the female was quite short at only about 1.3 m (4.3 ft) in height, much smaller than has been estimated for females prior to this discovery. Of greater significance is the shape of the pelvis, which indicates she could have given birth to an infant with a brain 30-50% the size of an adult’s. This suggests that the growth rate of the brain in the womb was similar to that of a modern human, but growth slowed in the first few years of life to a rate intermediate between modern humans and living chimpanzees.
In Asia, H. erectus lived 1.6 million years ago up to about 100,000 years ago with fossil remains recovered from sites in Indonesia, China, Turkey and the Republic of Georgia. Sangiran on the island of Java, is the most important Homo erectus site in Indonesia. The remains of over 80 individuals have been found here in a number of excavations. The site of Zhoukoudian, just south of Beijing in China, has yielded about 200 fossils from about 40 individuals, the largest number of H. erectus fossils from any one site. In addition, about 100,000 stone tool artifacts, mostly choppers and flakes of quartz and sandstone, were also in the cave. It was occupied between 200,000 and 750,000 years ago, although evidence suggests that occupation was sporadic rather than permanent, as hyenas and other animals also used the cave site. Other significant Asian H. erectus find include various fossils found at Dmanisi in the Republic of Georgia. At 1.8 million years old, these fossils currently represent the earliest known human presence outside of Africa. At the site of Kocabas, Turkey, skull fragments were found in a quarry that date between 500,000 and 400,000 years ago.
When compared with African H. erectus, Asian H. erectus had a robust skeleton that was generally shorter and stockier. The face was large with a low, sloping forehead, a massive brow ridge and a broad, flat nose, and the skull was broad and long with sharp angles at the rear. The bones of the skull were very thick and formed a small central ridge along the top of the skull, known as a midline keel. The lower jaw of Asian H. erectus was large and thick, and the molar teeth had large roots. Like African H. erectus, Asian H. erectus had limbs were like those of modern humans although the bones were thicker, suggesting a physically demanding lifestyle.
European H. erectus lived between 1.2 million and 400,000 years ago in Western Europe. Once again, there is some debate in taxonomic classification, with this group at times classified as Homo antecessor (1.2 million to 800,000 years ago) or Homo heidelbergensis (700,000 to 300,000 years ago). Partial remains of European H. erectus, associated stone tools, and animal bones were recovered from the cave site of Sima de Elefante in Atapuerca, Spain, all dating to about 1.2 million years ago. At a nearby site of Gran Dolina, remains of over 80 fossils representing at least 6 individuals, one of them a 10-year old child, and over 200 stone tools and 300 animal bones were found. While the human remains date to about 900,000 years ago, many of the stone tools and animal bones date to around 1.1 million years ago. Cut marks consistent with processing by humans for the purpose of obtaining meat and marrow are present on the animal remains at both sites. Interestingly, at Gran Dolina most of the human remains also display the same types of cut marks, indicating probable intentional dismemberment. There is also a lack of carnivore tooth marks on the human remains, supporting the suggestion that the remains were processed and not scavenged. Whether this was due to cannibalism is debatable, as there are instances of defleshing bones that do not involve consuming the flesh. However, these instances are usually related to funerary rites, and there is no evidence for this behavior in this species. Other significant H. erectus finds in Europe include a partial cranium from Ceprano, Italy dating to 800,000 years ago, a mandible with teeth from Mauer, Germany dating to 500,000 years ago, and a lower-leg bone from Boxgrove, England dating to about 500,000 years ago.
European H. erectus has a unique combination of features in the cranium, teeth and lower jaw that are collectively different from other Homo fossils, rather than any particular singular feature that distinguishes it from others. Body size and shape are similar to modern humans, but with a more robust build, especially in the legs. Adult males were about 1.7 m (5.6 ft) tall and weighed roughly 90 kg (200 lb). The skull has modern traits that include a projecting nose and hollowed cheekbones, but also archaic traits such as a low forehead, small post-orbital constriction, marked double arched brow ridge and a bulge of bone at the back of the skull known as an occipital bun. They also had robust teeth, but the lower jaw is thinner than that of African or Asian H. erectus, and for the first time we see some evidence of a receding chin.
By 500,000 years ago, H. erectus had become very adept at hunting large animals for food although the hides may also have been useful, especially in colder areas. The fossilized bones of these animals have shown that large animals including rhinos, hippopotamus, bears, horses and deer were targeted. These animals were skillfully hunted then butchered in an orderly fashion suggesting that these people were the earliest to work in co-operative groups. None of the H. erectus skeletons that have been found so far were deliberately buried. There is evidence however, that they did care for living members of their group who were sick or injured, but they did not seem to be concerned with their welfare after death.
Archaic Genus Homo
Evidence of H. erectus begins to disappear about 600,000 years ago and is replaced by a number of fossils within genus Homo that are grouped into the broad category of archaic humans. These individuals lived in Africa, Asia and Europe in the period contemporary to and predating the emergence of anatomically modern humans ( Homo sapiens ) 200,000 to 100,000 years ago. The archaic genus Homo grouping typically includes early archaic H. sapiens, H. naledi, H. neanderthalensis (Neandertals), H. denisova (Denisovans),
H. floresiensis, and H. luzonensis.
Early archaic Homo sapiens originated in Africa and lived from about 500,000 to 200,000 years ago in Africa, Asia and Europe (Figure 4.31). Between 600,000 and 200,000 years ago, the climates of Africa and Europe experienced a series of warm and cool phases and the move from Africa to Europe subjected these groups to generally colder climates. About 300,000 years ago, a severe cold, dry period began and the Sahara
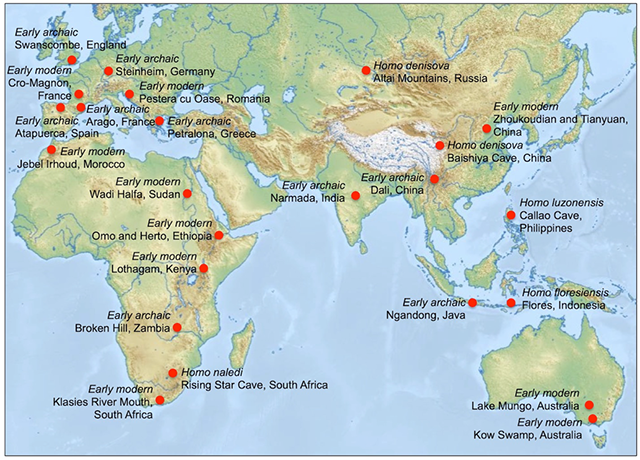
Figure 4.31: Sites of significant early archaic Homo sapiens and early modern Homo species fossil discoveries
became a barrier to movement between Africa and Eurasia, although movement may have been possible between Europe and northern Asia. At this time, populations in Africa and in Europe were isolated from one another and slight differences in skeletal traits began to appear due to regional adaptation. Individuals lived and worked in cooperative groups, hunted large animals and made a variety of tools including stone hand axes and wooden spears set with stone spearheads. Populations living in cooler European and Eurasian areas were most likely constructing animal hide clothing; however, direct evidence of clothing is lacking since it is tends to quickly degrade in the majority of the climates where early archaic H. sapiens have been found.
African and European fossils from this group are often taxonomically classified separately as H. rhodesiensis and H. heidelbergensis respectively rather than being identified as later H. erectus or early archaic H. sapiens. These fossils possess morphological features found in both H. erectus and modern H. sapiens. Common traits include a moderate double-arched brow ridge, sloping forehead, smaller teeth, wide nasal opening, long robust legs, and a notable expansion in brain size. Fossil evidence regarding body size and shape is currently limited but leg bones indicate they were tall, reaching about 1.8 m (5.9 ft) in height. Due to this mix of traits, most scholars recognize early archaic H. sapiens found in Europe and Africa as being close to the divergence of lineages that led to the Neandertals and early modern H. sapiens respectively.
Some of the most well known examples representing early archaic H. sapiens are from the Ngandong site on the island of Java, where multiple skulls have only the braincase preserved. These skulls have a long, low shape but having a larger brain case reflecting a change in brain size from earlier H. erectus fossils from the same region. At the Broken Hill site in Zambia, the skull of one individual showed significant tooth decay and a number of abscesses that are unusual for this time period, suggesting a great amount of sugars and starchy foods in their diet. The Arago Caves in Tautavel, France have revealed a number of human fossils including an intact skull of a young male with a wide face, thick arching brow ridges, sloping forehead and a large brain capacity. Thousands of stone tools and the bones of many types of animals have also been uncovered at this site.
Fossil remains of Homo naledi were discovered in 2013 in the Rising Star Cave system of South Africa. Although archaic features of its skeleton resembled fossil specimens roughly 2.0 million years old, these individuals date to 335,000 to 236,000 years ago, contemporary with Homo neanderthalensis, Homo denisova and anatomically modern Homo sapiens. Remains of H. naledi are an anatomical mosaic of traits similar to genus Australopithecus, mixed with those more characteristic of genus Homo, and traits that are not known in other hominin species. Adult males were about 1.5 m (5.0 ft) tall and weighed on average 45 kg (100 lb), with adult females slightly shorter and having less body mass. The shape of the pelvis is flared, similar to australopithecines, while the legs and feet are more similar to genus Homo. The hands are overall well suited for object manipulation, as they are more modern-like, but the fingers still retain a slight curvature and are more australopithecine-like. The arms, shoulders and ribcage are also similar in appearance to Australopithecus, but the vertebrae are very similar to genus Homo.
Skulls of H. naledi are closer in cranial volume to australopithecine skulls. However, the cranial structure is more similar to those of genus Homo, with slender features and no post-orbital constriction. The teeth are small, similar to early modern H. sapiens, but the
| Key Concepts: Archaic and Early Modern Genus Homo | |
|---|---|
|
Early archaic Homo sapiens ‘Atapuerca’ fossil |
Location: Africa, Asia, and Europe
Date(s): 500,000 to 200,000 years ago Habitat: Savanna, grassland, woodland and coastal environments Tool industry: Mode 2 (Acheulean) Key traits:
|

Homo naledi ‘Star’ fossil |
Location: South Africa
Date(s): 335,000 to 236,000 years ago Habitat: Limestone caves, grasslands and small forests Tool industry: Unknown Key traits:
|
|
Homo neanderthalensis ‘La Ferrassie’ fossil |
Location: Western Asia, Europe
Date(s): 200,000-27,000 years ago Habitat: Semi-glacial forests; similar to modern environments Tool industry: Mode 3 (Mousterian) Key traits:
|
| Key Concepts: Archaic and Early Modern Genus Homo | |
|
Homo denisova ‘Altai’ fossil |
Location: Russia and China
Date(s): X Habitat: Semi-glacial mountain steppes and forests Tool industry: Mode 3 (Mousterian) Key traits:
|

Homo floresiensis ‘Hobbit’ fossil |
Location: Flores, Indonesia
Date(s): 190,000 to 50,000 years ago Habitat: Tropical forest and mountain in island chain Tool industry: Unknown Key traits:
|
|
Homo luzonensis ‘Callao Man’ fossil |
Location: Callao Cave, Luzon, Philippines
Date(s): 67,000 to 50,000 years ago Habitat: Tropical forest and mountain in island chain Tool industry: Unknown Key traits:
|

Early modern Homo sapiens ‘Wise Man’ fossil |
Location: Africa, Asia, Europe, Oceania and the Americas
Date(s): 200,000 to 9,000 years ago Habitat: Environments similar to modern landscape Tool industry: Mode 4 (Aurignacian) and Mode 5 (Magdalenian) Key traits:
|
third molar is larger than the other molars, similar to australopithecines. The jaws are much smaller than those of most australopithecines, suggesting a diet that did not require heavy chewing. However, almost half of H. naledi teeth have chips on the enamel surface, caused by teeth coming into contact with plants with high silicate content, such as tubers and grasses, or environmental grit during life.
Attempts to extract DNA from H. naledi remains have thus far proven unsuccessful, but the H. naledi skeletons indicate that the origins of the genus Homo were complex and may be the result of hybridization, and that the species may have evolved separately in different parts of Africa.
The Neandertals appear in the fossil record about 200,000 years ago, with fossils exhibiting Neandertal-like characteristics appearing as early as 350,000 years ago. Evidence of Neandertals is found throughout Europe and into Central Asia, with the eastern-most occurrence of Neandertal fossils at the site of Teshik Tash, Uzbekistan (Figure 4.32). Some anthropologists classify Neandertals as a subspecies of H. sapiens, as H. sapiens neanderthalensis, while others interpret the morphological differences as significant enough to warrant classifying them as a different species, Homo neanderthalensis. Neandertals are recognizably human but have distinctive facial features and a stocky build that were evolutionary adaptations to cold, dry environments. Neandertals were generally shorter and had more robust skeletons and muscular bodies than early modern H. sapiens. Adult males were about 1.6 m (5.2 ft) tall and weighed on average 65 kg (143 lb). In comparison, adult females were about 1.5 m (5.1 ft) and weighed around 54 kg (119 lb). Their limbs were thick and had large joints, which indicates they had strongly muscled arms and legs, but limbs also tended to be shorter
Figure 4.32: Known extent of Neandertal world (blue) and sites with significant Neandertal fossil discoveries (Denisova Cave represents genetic evidence)
than those of early modern H. sapiens. These proportions are typical for people living in cold climates. They share a distinctive skull shape that was long and low, with a rounded brain case, an occipital bun and a depression for muscle attachments at the back of the skull. Facial features include a thick but rounded brow ridge under a relatively flat and receding forehead. The mid-facial region showed a characteristic forward projection from a large nasal opening and “inflated” sinuses. Their jaws were larger and more robust than those of early modern H. sapiens, and they still lacked a chin; however they had larger anterior teeth that on average exhibit a high level of wear from being used as tools.
It is commonly believed that Neandertals were the first early humans to wear clothing on a consistent basis, given the climate in which they were living. Tools such as stone scrapers and bone awls are often found associated with animal bones at Neandertal sites. Modern hunter-forager populations have historically used these kinds of tools for cleaning and processing animal hides. These hunter-forager groups also used their teeth as a “third hand” when lacing and constructing loose-fitting garments, weaving plant materials and softening small animal hides.
Neandertals were excellent hunters and are recognized as apex predators, regularly eating reindeer, wild boar and aurochs, and on occasion mammoth and rhinoceros. Neanderthals probably employed a risky and possibly less sophisticated hunting strategy than early modern Homo sapiens. There is a high frequency of healed bone fractures among Neandertal remains, which, along with their distribution in the body, are similar to injuries among professional rodeo riders who regularly interact with large, dangerous animals. The level of survival and healing after this type of major trauma indicates that they evidently cared for the injured and had some degree of medical knowledge.
Chemical analysis of their fossils shows that they ate significant amounts of meat, but also that their diet was wide-ranging and supplemented with nuts, mushrooms and vegetation. Despite this mixed diet, nearly half of Neandertal skeletons show the effects of a diet deficient in nutrients. It has long been debated whether Neandertals also included human meat in their diets. It is not always easy to determine if cut marks on human bones are due to cannibalism or some other practice, but in recent years new evidence has emerged that suggests some Neandertals may indeed have been cannibals on occasion. Bones from Abri Moula in France show cut marks typical of butchery rather than simple ritual defleshing. The marks were also like those on the bones of roe deer, assumed to be food, found in the same shelter. El Sidrón Cave in Spain yielded hundreds of Neandertal bones with cut marks, deliberate breaks for marrow extraction, and other signs that the bodies had been butchered for flesh in the same way as animals.
A significant number of Neandertal fossils in Europe and Western Asia have been found in features described as burials. These pits were purposefully dug with the corpse placed centrally and then filled in. Hands and arms are positioned and the majority of the bodies were on their sides or backs. In some instances, other objects, such as animal bones or tools, or traces of vegetation have been found with the body. This kind of treatment suggests the bodies were not randomly disposed of, but instead represent some kind of symbolic or personal link between the living and the dead. Whether Neanderthals created art and used personal or group adornments, which would indicate a capability for complex symbolic thought, remains unresolved; however, in 2018, red painted symbols, negative hand imprints and red lines and dots on the walls of three Spanish caves were dated to 64,000 years ago. In addition, researchers found perforated seashell beads and pigments at least 115,000 years old in in Cueva de los Aviones, Spain. Both finds date to before early modern H. sapiens are thought to have arrived in Europe.
The current fossil record from France and Spain indicates that Neandertals went extinct between 35,000 and 27,000 years ago. Various reasons have been proposed for early modern H. sapiens replacing Neandertals, often suggesting that these “sluggish brutes” stood no chance against the “vastly superior” humans . Today, a majority of scientists accept that Neandertals displayed advanced social and cultural behaviors and complex adaptive strategies, but incoming early modern H. sapiens appear to have had subtle differences in biology and behavior that gave them an edge under the social circumstances and changing climatic conditions.
While we are closely related to the Neandertals, they are not our direct ancestors. Evidence from the fossil record and genetic data shows they are a distinct species that developed as a side branch in our family tree. The Neandertal genome shows that early modern H. sapiens and Neandertals did interbreed, although on a very limited scale. Those of European and Asian descent share about 1-4% of their nuclear DNA with Neandertals, while those of African descent share no nuclear DNA with Neandertals. This suggests that early modern H. sapiens bred with Neandertals after early moderns left Africa but before they spread to Asia and Europe. The most likely location is the Levant, where both species co-existed at various times between 90,000 to 50,000 years ago. Other key genetic findings show the presence of the MC1R gene for red hair and fair skin and the FOXP2 gene related to language ability the same as modern humans.
The Denisovans, pending widely recognized taxonomic status, currently carries the temporary species Homo denisova . Remains representing the Denisovans include were several teeth, long bone fragments and a small fragment of finger-bone dated between 195,000 to 43,000 years ago and found in Denisova Cave in the Altai Mountains of Siberia, Russia, and more recently a lower jaw fragment with teeth dating to 40,000 to 30,000 years ago from Baishiya Cave in Tibet (China) (Figure 4.31). Analysis of the jaw fragment shows a variant of the EPAS1 gene that reduces the amount of oxygen-carrying protein hemoglobin in their blood, enabling them to live at high altitudes with low oxygen levels.
Archaeological evidence of culture among Denisovans is so far limited. Stone tools were apparently not much different from other Paleolithic populations in the Siberian north. Decorative objects of bone, mammoth tusk, and fossilized ostrich shell were recovered from Denisova Cave, along with fragments of a stone bracelet and an eyed bone needle.
Genetic evidence suggests the Denisovans were once widespread across Eurasia and interbred with both Neandertals and early modern H. sapiens. The mitochondrial DNA of a fragment of finger bone from Denisova Cave was significantly different from both Neandertals and early modern H. sapiens, revealing that the Denisovans were a sister group to Neandertals, both descending from a population that split away from early modern H. sapiens about 765,000 to 550,000 years ago. At Denisova Cave some 80,000 years ago, Neandertals and Denisovans reconnected. DNA evidence from one long bone fragment points to a hybrid individual who had a Denisovan father and a Neandertal mother. While Neandertal DNA is present in all populations outside of Africa, Denisovan DNA is only found in modern populations from China, island Southeast Asia, and Oceania.
Homo floresiensis lived from about 190,000 to 50,000 years ago in the Indonesian islands (Figure 4.31). The unfossilized remains include a largely complete skeleton with skull and parts of at least eleven other individuals recovered from Ling Bua Cave (Figure 4.33) on the island of Flores. The species disappearance coincides with that of other local fauna such as the pygmy elephant Stegodon and the giant carnivorous marabou stork. The arrival of early modern H. sapiens quite possibly played a role in their demise. Although there is no evidence of early modern H. sapiens in Liang Bua Cave until 11,000 years ago, other archaeological evidence indicated they were moving through the region about 50,000 years ago.
Figure 4.33: Entrance and excavations in Liang Bua Cave, Flores, Indonesia
Stone tools have been recovered from a number of levels within the cave and range in dates from 190,000 to 50,000 years old. Analysis of the residues and polish on some of the tools revealed they were used for working wood and fibrous materials, perhaps to make spear shafts or items such as traps. Cut marks on the Stegodon bones also suggest some of the tools were used to process meat. There is evidence of the use of fire in Liang Bua cave. The remains of numerous juvenile Stegodon have charred bones, possibly indicating that H. floresiensis was able to control fire for cooking.
The unusual features and recent survival of H. floresiensis suggests our human family tree is more complex than once thought. When first discovered, it was suggested that H. floresiensis was possibly descended from Javanese H. erectus. However, more detailed analysis of skeletal remains has uncovered traits more archaic than Asian H. erectus and more similar to australopithecines and H. habilis. It is now suggested that its ancestor may have come from an early African dispersal by an ancestral Homo species similar in appearance to H. habilis or Dmanisi H. erectus. Stone tools dating to 840,000 years ago were discovered on Flores, indicating that H. erectus or a similar species of hominin was probably living on the island at that time.
The most complete skeleton is that of a female aged about 30 years old. She stood about 1 m (3.2 ft) tall, weighed about 25 kg (55 lb) and had a small cranial capacity. Her body was not deliberately buried but covered soon after death by fine cave sediments. This species underwent insular dwarfism, or reduction in size over a number of generations due to long term isolation on the island, which resulted in an endemic ‘dwarf’ population. This is a common phenomenon seen in other mammals in this environment. They had a long and low cranial shape, closer to that of H. erectus than H. sapiens, and a brain that includes features that are not found in H. sapiens or earlier genus Homo species. The lower jaw had a V-shaped dental arcade typical of Homo, but there was also a bony shelf at the front of the lower jaw which is an ancestral feature not seen in genus Homo. H. floresiensis had long arms, a wide pelvis, and a relatively long foot for its body size (70% as long as the thigh bone, compared with 55% for early modern H. sapiens ), a flat arch lacking the spring-like mechanism used to store and release energy during running, and a short big toe. These features are similar to ancient hominins such as H. habilis.
Homo luzonensis has recently been assigned to genus Homo as a new species based on a small number of specimens from the island of Luzon in the Philippines (Figure 4.31), dated 67,000 to 50,000 years ago. However, some scientists think additional evidence is required to confirm the fossils as a new species, rather than a locally adapted population of other genus Homo populations, such as H. erectus, as the fossils present a mosaic of earlier and modern traits. To date, H. luzonensis is represented by 10 small hand and foot bones, 14 teeth and a femur shaft that represent at least three individuals. The small sizes of the molars suggest that it may have undergone island dwarfing similar to H. floresiensis, although no estimate of its height is currently possible. Curvature in the foot bones is similar to Australopithecus, indicating these individuals may have retained some level of arboreal adaptation. The fossil human remains were associated with animal bones exhibiting potential cut marks, suggesting that they were butchered; however, no stone tools dating to the same period have been recovered as yet. In 2016, researchers uncovered stone tools on the island of Sulawesi dating to 150,000 years ago, at least 60,000 years older than the island’s oldest known early modern H. sapiens. Taken alongside the remains from the islands of Flores and Luzon, these Indonesian and Philippine sites suggest that ancient hominin dispersal throughout the region wasn’t necessarily as rare or as accidental as researchers once thought.
Early Modern Genus Homo
African fossils provide the best evidence for the evolutionary transition from early archaic to early modern Homo sapiens around 200,000 years ago. This group is also often referred to as anatomically modern H. sapiens and is found in Asia by 100,000 years ago and in Europe by 35,000 years ago (Figure 4.31). Within the last 20,000 years early modern H. sapiens also spread from Asia into Oceania and the Americas.
The earliest modern H. sapiens had bodies with short, slender trunks and long limbs. These body proportions were an adaptation for surviving in tropical and sub-tropical regions. More stocky builds gradually evolved when populations spread and adapted to cooler regions. Adult males were about 1.6 m (5.2 ft) tall and weighed on average 60 kg (132 lb). In comparison, adult females were about 1.5 m (5.1 ft) and weighed around 50 kg (110 lb). The limbs are thinner and less robust than earlier archaic species, indicating a clear reduction in muscle size, with the legs relatively long when compared with the arms. The pelvis is narrow from side-to-side but a deeper bowl-like appearance from front-to-back. Bones in the hands are straight and capable of very fine precision grip and manipulation of objects.
Homo sapiens living today have an average brain size of about 1350 cc which makes-up 2.2% of our body weight. Early modern Homo sapiens, however, had slightly larger brains at nearly 1500 cc. This is reflected in changes to the shape of the skull, which has a short base and a high braincase. Unlike other species of genus Homo, the skull is broadest at the top. The fuller braincase also results in almost no post-orbital constriction. The back of the skull is rounded, with a reduction in muscle attachment structures (e.g., nuchal crest or occipital bun). The face is small with a projecting nose, square-shaped eye orbits, a smaller brow ridge and a tall forehead. The jaws are shorter, which results in a vertical, flat appearance, and slight in build with a bony chin projection for added strength. Homo sapiens is the only species with a protruding chin. The teeth are relatively small when compared with early archaic H. sapiens and are now arranged in a parabolic shape with side rows further apart at the rear of the mouth, forming a clear V-shaped pattern.
Early modern H. sapiens had a more complex culture and technology than what is currently recognized in previous species. Evidence for symbolic behavior appears at a number of South African sites about 125,000 to 100,000 years ago. Red ochre pieces from Blombos Cave in South Africa, dating to about 100,000 to 80,000 years ago, show evidence of engraving that may be an expression of art or markings made during other activities. However, other signs of possible symbolic behavior, including shell beads and sophisticated stone tools have also come from this site, strengthening the case for early artistic expression.
The archaeological record across Africa, Europe and Asia has produced an explosion of artistic material from around 40,000 years ago. Many researchers believe this shift is tied to a change in human cognition and communication. Population size and structure are thought to have played key roles, as social learning is considered more beneficial to developing complex culture than are individual innovations. The cave art being produced about 40,000 years ago depicts animals (Figure 4.34) or probable spiritual beings, but smaller marks in many caves in France, and possibly others in Europe, are now being analyzed as a written ‘code’ familiar to many early modern H. sapiens groups. There are over 20 symbols that appear repeatedly across thousand of years, some of them in pairs and groups in what could be a rudimentary ‘language,’ suggesting that early modern H. sapiens in Europe were attempting to represent ideas symbolically rather than realistically and share information across generations.
Figure 4.34: Cave painting of animals at Chauvet Cave, France
Initially, early modern H. sapiens made stone tools that were similar in design to those made by the Neanderthals. It required the ability for abstract thought to mentally plan a series of steps that could then be executed in tool production. As more sophisticated techniques developed in some parts of the world, this early technology was soon replaced with the use of a wider range of materials including bone, ivory and antler. Evidence of musical instruments first appeared about 32,000 years ago in Europe in the form of bone flutes and whistles from various sites in France.
Sophisticated control of fire, including complex hearths, pits and kilns, allowed early modern H. sapiens to survive in regions that even the cold-adapted Neanderthals had been unable to inhabit. They often inhabited caves or rock shelters if these were available. More recently, especially within the last 20,000 years, natural shelters were enhanced with walls or other simple modifications. In open areas, shelters were constructed using a range of framework materials including wooden poles and the bones of large animals, such as mammoths. These structures were probably covered with animal hides and the living areas included fire hearths. The site of Dolni Vestonice in the Czech Republic produced the earliest evidence for high temperature kilns and ceramic technology. The kilns, dated at 26,000 years old, were capable of firing clay figurines at temperatures over 400 °C (752 °F). About 2000 fired lumps of clay were found scattered around the kiln.
Animal hide clothing may have been worn in cooler areas, although direct evidence of clothing only exists for the last 30,000 years, including specialized tools such as eye needles, adornments such as buttons and beads sewn onto clothing, and the remains of animals such as arctic foxes and wolves that indicate they were trapped for their fur. Clothes that were sewn provided better protection from the cold than clothes that were laced together. Flax fibers, discovered in a cave in the Republic of Georgia and dating to about 36,000 years ago, were most likely used to make clothes and woven baskets, and a small number appear to be dyed. Textile impressions have been discovered at other European sites have, but no actual remains, making this find the earliest of its kind.
Items of personal adornment not sewn onto clothing include ivory, shell, amber, bone and tooth beads and pendants. Ostrich eggshell beads that date from about 45,000 years ago have been found in Africa, as well as pierced shell beads in Morocco dating to 80,000 years ago and marine shell beads from Israel dating to 90,000 years old, but evidence of body adornment become prolific by about 35,000 years ago. One of the earliest known pendants is a horse carved in mammoth ivory around 32,000 years ago in Germany.
Burials were infrequent and fairly simple prior to 40,000 years ago and then began to become more elaborate with the inclusion of valued objects such as tools and body adornments. Red ochre was sprinkled over many of the bodies prior to burial. One of the earliest deliberate burials of a modern human comes from Jebel Qafzeh in Israel. Dating to 90,000 years ago, the grave contains the bones of a young woman buried with a young child at her feet. An additional 21 skeletons were also found in the same cave.
Tool Industries
Stone tool technology refers to a broad array of techniques and styles to produce usable tools from various types of stone. Stone tools and other artifacts offer evidence about how early humans made things, how they lived, how they interacted with their surroundings and evolved over time.
Earliest Tool Industries
Pre-Mode 1 industry, also referred to as Lomekwian, is based on stone tools found at Lake Turkana, Kenya that date to 3.3 million years ago, suggesting that they may have been produced by Australopithecus species. The tools include large, modified chipped cores (Figure 4.35), which are the source material for stone tool production, and flakes, or thin fragments of stone removed from a core. Analysis suggested the cores had been rotated as flakes were struck off. This is the earliest recognizable tool technology preserved in the archaeological record, and the tools seem to be a simple progression from the use of naturally modified stones that our earliest ancestors probably used.
Figure 4.35: A Pre-Mode 1 Lomekwian industry stone core
Mode 1 industry was a widespread stone tool technology of the Lower Paleolithic period used from 2.6 to 1.7 million years ago across much of Africa, South Asia, the Middle East and Europe. Tools from this industry are often referred to as Oldowan, as the first discoveries of these tools occurred in Olduvai Gorge, Tanzania in East Africa.
To obtain a Mode 1 stone tool, a spherical hammerstone is used to strike the edge of a core, in order to fracture it and remove flakes. When the core has an edge on one side and can be used as a tool, it is referred to as a chopper (Figure 4.36). The earliest chopper tools were simply split river cobbles made of quartzite, obsidian, and chert. These kinds of stone, when fractured, can produce a very sharp edge, which can be used for scraping, chopping, cutting and boring holes.
Figure 4.36: A Mode 1 Oldowan industry stone chopper tool
Distinct Tool Technologies
Mode 2 industry dates to the Lower Paleolithic from 1.7 million years ago up through the early Middle Paleolithic until about 160,000 years ago and was used across Africa, Europe, and much of West Asia and South Asia. Tools from this industry were first discovered during the 1800s at St. Acheul in France, which is why they are more commonly referred to as Acheulean.
This new technology was developed by H. erectus in Africa and was an improvement on the very simple stone choppers that earlier ancestors such as H. habilis had been using for about 1.0 million years. In Europe and Western Asia, early Neanderthals adopted Acheulean technology, transitioning to a newer industry by about 160,000 years ago. Unlike the earlier Mode 1 industry, the core was prized over the flakes that came from it in Mode 2. Some smaller tools were made from large flakes that had been struck from stone cores. These flake tools and the distinctive waste flakes produced in manufacture suggest a more considered technique, one that required the toolmaker to think one or two steps ahead during work that necessitated a clear sequence of steps to create perhaps several tools in one sitting.
Another advance was that the Mode 2 tools were worked symmetrically and on both sides indicating greater care in the production of the final tool. This industry is characterized by distinctive oval and pear-shaped bifacial hand-axes, but also includes straight edge cleavers and scrapers (Figure 4.37). These tools were suitable for heavy-duty work including butchering large mammals, processing bones for marrow, and working other materials such as bone, antler and wood.

Figure 4.37: A Mode 2 Acheulean industry stone bifacial hand axe tool
A hard hammerstone would first be used to rough out the shape of the tool from the stone by removing large flakes. The toolmaker would work around the circumference of the remaining stone core, removing smaller flakes alternately from each face. The scar created by the removal of the preceding flake would provide a striking platform for the removal of the next. Once the rough shape was created, a further phase of flaking was undertaken to make the tool thinner. The thinning flakes were removed using a softer hammer, such as bone or antler. Final shaping was then applied to the usable cutting edge of the tool, again using bone, wood or antler for fine removal of flakes.
New Materials, New Technologies
Eventually, the Mode 2 industry in Europe was replaced by a lithic technology known as the Mode 3 industry, or Mousterian, which was named after the site of Le Moustier in France, where examples were first uncovered in the 1860s. Mode 3 Industry was developed and used primarily by the Neandertals in Europe and Western Asia, but a broadly similar contemporaneous industry was in widespread use in Africa.
This industry adopted the Levallois technique to produce smaller and sharper knife-like tools called blades as well as small bifaces, scrapers and points (Figure 4.38). Also known as the “prepared core technique,” flakes are struck from worked cores and then subsequently retouched.
As more sophisticated techniques developed in some parts of the world, this early Mode 3 technology was replaced by either Mode 4 or Mode 5 industries and the use of a wider range of raw materials including bone, ivory and antler.
Figure 4.38: A Mode 3 Mousterian industry tool kit of stone points, scrapers, blades, and small bifaces
Mode 4 industry, developed by Aurignacian culture groups, first appeared in Africa during the Upper Paleolithic and was used between 50,000 and 10,000 years ago. It is characterized by the production of long, thin stone flakes shaped into blade knives, spearheads and other composite material tools. The production of these long, thin blades as a base tool type exponentially increased the efficiency of core usage compared to the Levallois prepared core flake technique, which had a similar advantage over Acheulean industry, which was directly worked from cores.
Mode 5 industry, used by Magdalenian culture groups, specialized in the production of very small blades called microliths that were mainly fastened to a shaft of composite tools having several parts. These tools included small-headed arrows, barbed spears and sickles. Regional variation in these tool cultures developed with an influx of new styles, techniques and cultural needs, especially within the last 13,000 years as domestication of plants and animals started to occur independently in various locations around the world.
Models of Human Origins
There are three models widely used to interpret the origin of living human populations: the Replacement, or ‘Out of Africa’ model; the Continuity, or ‘Multiregional Evolution’ model; and the Assimilation, or “Hybrid’ model.
The Replacement model posits that all living Homo sapiens descend from a population that first appeared in Africa and subsequently spread throughout that continent and across the remainder of the globe, replacing existing hominin populations such as the Neanderthals. This would have occurred sometime between perhaps 90,000 and 60,000 years ago.
Proponents of the Continuity model suggest that hominin populations have maintained extensive social and biological ties across Africa and Eurasia for at least the last 1.8 million years, beginning when members of ancestral species such as Homo erectus dispersed from Africa. In this model, isolation among hominin populations created regional variation in behavior and biology, but occasional contact between groups through the sharing of ideas and mating (gene flow) was sufficient to prevent the formation of distinct biological species.
The Assimilation model combines elements of the other two models, and adherents of this model favor an African origin of Homo sapiens with varying degrees of integration through interbreeding with existing populations as Homo sapiens spread from their place
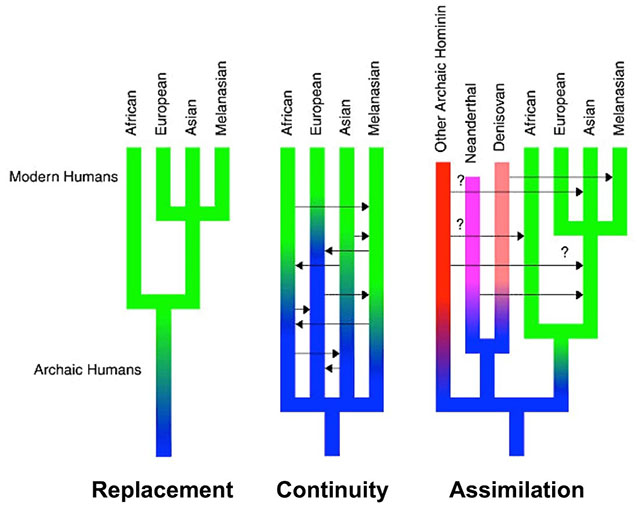
Figure 4.39: Models used to interpret the origin of living human populations
of origin. This is commonly seen as a more flexible version of the Replacement model, where archaic Homo sapiens are leaving Africa and not just “replacing” existing populations but also mating with them, creating hybrids.
Evidence needed to support or refute these models include the age and location of the earliest fossils assigned to Homo sapiens, the species attribution and fate of pre-existing hominin populations (e.g., the Neandertals), and the extent to which modern and ancient DNA evidence for interbreeding can be found in the fossil record.
Some paleoanthropologists add an additional complication: Did anatomical modernity and behavioral modernity appear separately, or did they arise simultaneously? According to one view, modern human form preceded the appearance of what we would recognize as modern cognitive abilities. These abilities include the extensive use of symbols, both verbally, as fully developed syntactical language, and visually, as art and personal adornment, as well as the development of complex extended social networks. Evidence suggest that behavioral modernity arose as an advantageous genetic mutation resulting in neural reorganization that spread rapidly among existing populations of anatomically modern humans perhaps 50,000 years ago. The alternative view is that the capacity for modern human behavior is coincident with the origins of Homo sapiens, and that the expression of this capacity in the archaeological record was an incremental process developed through time. The data needed to test these models includes artifacts and other elements of the archaeological record; in other words, the material traces of past human behavior. These include objects and tools of bone, stone, antler, and shell, evidence for how they were made and used, and features such as hearths, that show control of fire and modification of the environment
Review of Learning Objectives
Careful comparison between humans and other primate species offers enormous insight into our evolutionary past. Primatologists attempt to make sense of primate diversity by creating a primate taxonomy. Strepsirrhines include lemurs , lorises and galagos. Haplorhines include tarsiers and anthropoids. Anthropoids include New World and Old World forms. New World monkeys differ from Old World monkeys and apes in nose shape, dentition, and prehensility. All New World monkey species are arboreal. Old World anthropoids include monkeys and apes, as well as humans. Apes are distinguished fro Old World monkeys by dentition, skeletal shape and size, and the absence of a tail. The African apes are far more closely related to one another than they are to gibbons or orangutans, and humans are more closely related to chimpanzees and bonobos than to any other ape species.
Paleoanthropologists study the origins and development of early humans and their ancestors by focusing on the fossil record. Bipedalism, reduction in tooth size, expansion of cranial capacity and the development of tool technology are all a part of the mosaic of evolutionary trends seen in our ancestors. Hominins were for the most part successful in exploiting food recourses on the ground, outside the protection of forests. Their diet was probably omnivorous, and they could move efficiently in groups while carrying infants, food, and tools.
The earliest bipedal hominin skeletal fossils of Sahelanthropus are approximately 7.0 million years old, followed by a few transitional species in genus Ardipithecus, but by 3.0 million years ago, hominin diversity increased to included several species from genus Australopithecus and Paranthropus. These individuals displayed dental and skeletal traits that indicate specialized adaptations to niche environments in Africa and possibly evidence of the earliest use of stone tools. The first members of the genus Homo appeared about 2.5 million years ago, coexisting with Australopithecines for a short time. Around 1.8 million years ago, Homo erectus left Africa and migrated into Europe and Asia. Homo erectus had a diet different from previous hominins, which included a greater amount of protein obtained from scavenged and hunted animals. Their skulls show an increase in cranial capacity, and the archaeological record contains evidence of the first tool industry and the first controlled use of fire. Between 500,000 and 200,000 years ago Homo erectus begins to disappear and archaic Homo sapiens and other regionalized species take their place across the landscapes of Africa, Europe and Asia. This transition was accompanied by anatomical adaptations to regional environments, including an expansion of cranial capacity equivalent to modern Homo sapiens. By 80,000 years ago, stone tool traditions, art forms, symbolism, and lifeways were much more complex. As these archaic and early modern ancestors regularly hunted large game, constructed shelters, and expressed themselves with personal ornamentation and portable art objects, we see the most striking evidence for the modern human capacity for culture.
Concept review
- List the primary characteristics and forms of locomotion seen in primates.
- Summarize the features used to distinguish New World and Old World primates from each other.
- Explain why and how humans are taxonomically a part of the order Primates.
- Distinguish the difference between the terms hominid and hominin.
- Define bipedalism and explain its importance in human evolution.
- Explain the differences between species in the genera Australopithecus and Paranthropus.
- Summarize what is known about the morphology and behavior of Homo erectus.
- Describe the lifeways of Neandertals and early modern Homo sapiens.
- List the features used to emphasize and distinguish anatomically modern Homo sapiens from other archaic populations of genus Homo.
- Describe the increasing technological complexity in tool production from the Pre-Mode 1 through Mode 5 industries.
- Compare and contrast the three main models of modern human origins.
Applying concepts
- Discuss the ways in which evolutionary forces might contribute to the vast anatomical and dietary diversity seen in the order Primates.
- Observe a group of humans in a public place and compare their traits with traits that are distinctive in apes. Write short essay that explains your findings.
- Use Internet resources to explore the social behavior, ecology, and conservation status of one species of New World or Old World monkey. Write a paragraph that reports the information.
- Make a chart of the mosaic of trends seen in the dentition, pelvis and limbs from Australopithecus through archaic genus Homo.
- Use Internet resources to explore the various explanations for the disappearance of Neandertals. Describe the evidence that supports at least two of these explanations.
- Discuss evidence that supports our genus Homo lineage as being more similar to a “family bush” than a “family tree.”
