
2
Integrating Evolution and Genetics

Learning Objectives:
- Briefly outline evidence for evolution and deep geologic time
- Explain how the modern synthesis of evolutionary theory is based on earlier scientific thought on evolution and heredity
- Summarize principles of inheritance and understanding of variation with extended knowledge of molecular genetics
- Describe the mechanisms and processes of evolutionary change and occurrence of speciation within populations
Theories of Evolution
All species of living organisms evolved at some point from a different species. Although it may seem that living things today stay much the same from generation to generation, that is not the case: evolution is ongoing. Evolution is simply the process of change over time. Today, with the modern technology of molecular genetics, we are able to more specifically define evolution as the process of change in the heritable characteristics of biological populations over successive generations.
The theory of evolution is the unifying theory of biological sciences. The principle that all life has evolved and diversified from a common ancestor is the foundation from which biologists derive questions about the natural world. Since biological anthropologists study the relationship between biology and culture, this unifying theory and principle serve as a framework within which biological anthropologists study the living world and our past.
The theory of evolution by natural selection describes a mechanism for species change over time. That species change had been suggested and debated well before Charles Darwin. The view that species were immutable, or static and unchanging, was grounded in the writings of the ancient Greek philosopher Plato, but there were also a number of ancient Greeks that came before him and proposed that one type of organism could descend from another type.
In the eighteenth century, ideas about the evolution of animals were reintroduced by the naturalist Georges-Louis Leclerc, Comte de Buffon and even by Darwin’s grandfather, Erasmus Darwin. During this time, it was also accepted that there were extinct species. At the same time, James Hutton, the Scottish naturalist, proposed the theory of uniformitarianism, or the concept that changes in the earth’s crust over deep geological history have resulted from the action of continuous and uniform processes, such as erosion, volcanic eruptions and plate tectonics. This contrasted with the ideas of the French naturalist, Georges Cuvier, who proposed that the geology of the planet and the fossilized animals it contained were a consequence of catastrophic events occurring during a relatively brief past, a perspective known as catastrophism. Later in the nineteenth century, the geologist Charles Lyell popularized Hutton’s view of uniformitarianism. Lyell was a friend to Darwin and his ideas were very influential on Darwin’s thinking. Lyell argued that the greater age of Earth gave more time for gradual change in its geology, and the process provided Darwin with an analogy for gradual change in species.
In 1809, Jean-Baptiste Lamarck published a book detailing a mechanism for evolutionary change that is now referred to as inheritance of acquired characteristics. In Lamarck’s theory, modifications in an individual caused by its environment, or the use or disuse of a structure during its lifetime, could be inherited by its offspring and, therefore, bring about change in a species (Figure 2.1). While this form of inheritance and mechanism of evolutionary change as described by Lamarck was discredited, Lamarck’s ideas were and still are an important influence on evolutionary thought.
Figure 2.1: Lamarckian concept of inheritance through use over time; original short-necked giraffe ancestors continually stretch their necks to reach higher leaves, and over time, their necks becomes progressively longer in descendent generations
Keys to evolutionary theory
Six scientific disciplines have played key roles in the development of the theory of evolution by natural selection: taxonomy, biological systematics, geology, paleontology, evolutionary biology and demography.
Taxonomy is the science of defining and naming groups of biological organisms on the basis of shared characteristics. Organisms are grouped together into taxa, and these groups are given a taxonomic rank, thus creating a taxonomic hierarchy (Figure 2.2).
In the eighteenth century, the Swedish naturalist Carl von Linné (Latinized as Carolus Linnaeus) developed a system known as Linnaean taxonomy for categorizing organisms and binomial nomenclature for naming organisms. The first part of the name – the generic name – identifies the genus to which the species belongs, while the second part – the specific name – identifies the species within the genus. For example, humans belong to the genus Homo and within this genus to the species Homo sapiens. Tyrannosaurus rex is probably one of the most widely known binomial nomenclatures. Through careful comparison and analysis, von Linné and those who have come after him have been able to classify specific animals into a series of larger and more inclusive groups.
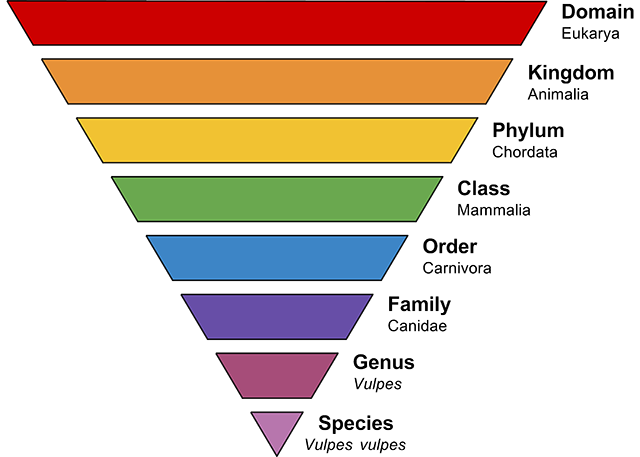
Figure 2.2: Taxonomic ranking designates related animals through main taxonomic ranks, such as domain, kingdom, phylum, class, order, family, genus, and species.
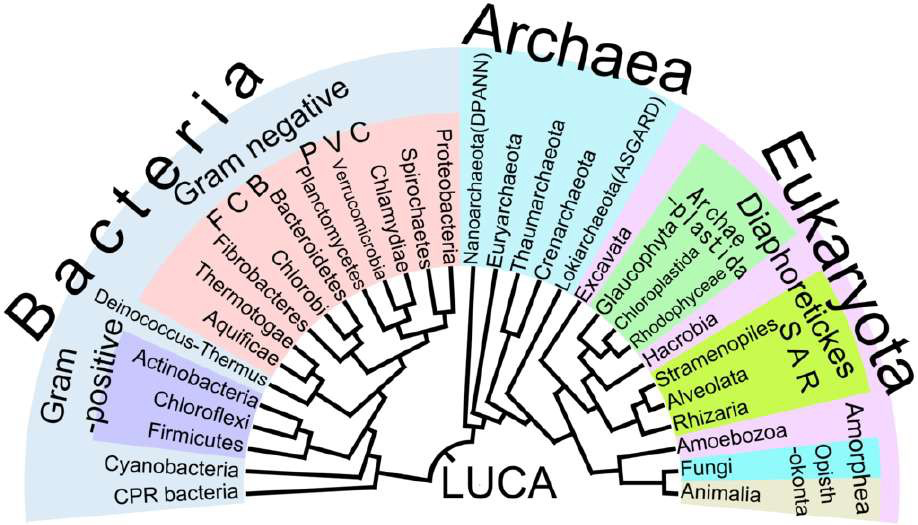
Figure 2.3: Phylogenetic tree of life branching diagram which shows the inferred relationship between various biological species. The distance of the branches in the phylogenetic tree represents the amount of inferred evolutionary change from the last universal common ancestor (LUCA).
Biological systematics is the study of diversification of living forms, both past and present, and relationships among living things through time. Relationships are visualized as evolutionary trees. Phylogenetic trees of species and higher taxa (Figure 2.3) are used to study the evolution of traits, such as anatomical or molecular characteristics, and biogeography, or the distribution of species and ecosystems in geographic space and through geological time.
Geology is the study of the structures of the Earth on and beneath its surface, and the processes that have shaped those structures. It also provides tools to determine the relative and absolute ages of rocks found in a given location, and also to describe the histories of those rocks. By combining these tools, geologists are able to chronicle the geological history of the Earth as a whole, and also to demonstrate the age of the Earth. Geology provides the primary evidence for plate tectonics, the evolutionary history of life, and the Earth’s past climates.
Paleontology is the study of ancient plant and animal life. It includes the study of fossils to determine organisms’ evolution and interactions with each other and their environments. Paleontology seeks to map out how living things have changed through time. The geologic time scale (Figure 2.4) is the “calendar” for events in Earth history. Paleontologists record major changes and events in ancient plant and animal life in a series of eons, eras, periods, and epochs, all of which are assigned numerical ages, usually with the notation of Ga, for billions of years ago and Ma for millions of years ago.
Fossil evidence indicates that during the Archean eon, which dates from 4.0 billion to 2.4 billion years ago, simple forms of life resembling modern bacteria emerged, some of which were able to photosynthesize. During the Proterozoic Eon, from 2.5 billion to 541 million years ago, the dominant life forms were algae, fungi and marine protozoans.
The Paleozoic Era, which dates from 541 million to 252 million years ago, was a time of dramatic geological, climatic, and evolutionary change. The start of this era witnessed the most rapid and widespread diversification of life in Earth’s history in an event known as the Cambrian Period explosion. Life began in the ocean but eventually transitioned onto land, and by the late Paleozoic, it was dominated by various forms of organisms including fish, amphibians, reptiles, and the first vertebrate land mammals. Great forests of primitive plants covered the continents. The Paleozoic ended with the largest extinction event in the history of Earth, the Permian-Triassic extinction. The effects of this catastrophe were so devastating that it took life on land 30 million years into the Mesozoic Era to recover.
The Mesozoic Era, dating to 252 to 65 million years ago, is known as the Age of Reptiles. It is divided into the Triassic, Jurassic, and Cretaceous periods. Dinosaurs first appeared in the Early Jurassic, and became the dominant terrestrial vertebrates for 135 million years before their demise at the end of the Cretaceous. Birds, having evolved from a branch of dinosaurs, first appeared in the Jurassic, and the first small mammals appeared in the Cretaceous. The Mesozoic ended with the Cretaceous-Paleogene extinction event, where changing climatic conditions, competition from newly evolving mammals, and a possible meteor impact with Earth, resulted in approximately 75% of all life over 10 kilograms becoming extinct, including non-avian dinosaurs.
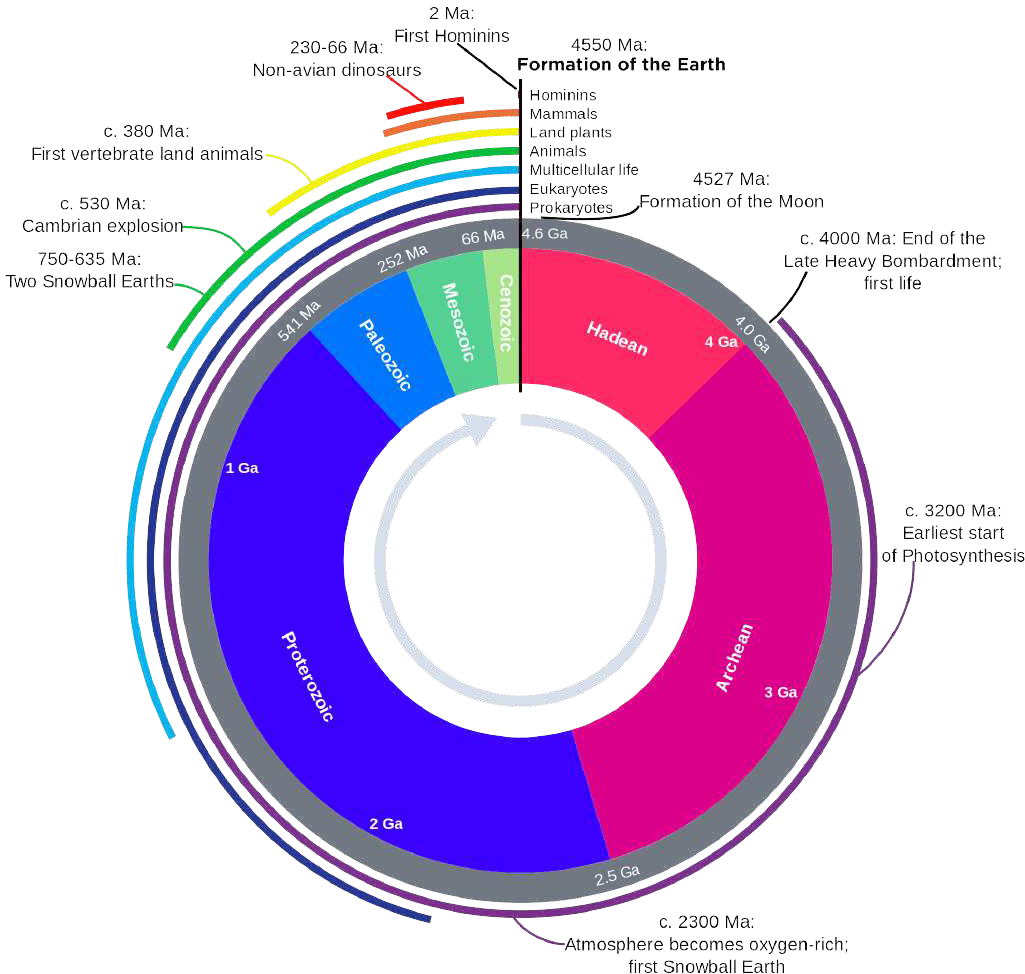
Figure 2.4: This clock representation shows some of the major units of geologic time and definitive events of Earth history. The Hadean Eon represents the time before fossil record of life on Earth, while the other subdivisions reflect the evolution of life.
The Cenozoic Era, also known as the Age of Mammals, extends from 65 million years ago to the present. Mammals came to occupy almost every available niche (both marine and terrestrial), and some also grew very large, attaining sizes not seen in most of today’s mammals. The Cenozoic is divided into three periods, the Paleogene, which encompasses the first 42 million years, and the Neogene, which covers the next 20.5 million years, and the Quaternary, which covers the last 2.5 million years. The earliest known primates appeared in the Paleogene during the Eocene Epoch between 56 million and 34 million years ago. We see the emergence of the genus Homo during the Neogene in the Pliocene Epoch from 5 million to 2.5 million years ago. The Quaternary Period is divided into two epochs. The Pleistocene Epoch, which extends from 2.5 million to 11,700 years ago, is marked by four separate ice ages to the north and severe drought that created the African desert regions. The second epoch, the Holocene, began around 11,700 years ago and lasts to the present day. All recorded history modern humanity lies within the boundaries of the Holocene epoch.
Evolutionary biology studies the evolutionary processes that produced the diversity of life on Earth, starting from a single common ancestor. These processes include natural selection, common descent, and speciation. The discipline emerged during the period of the Modern Synthesis in the 1930s and 1940s when ideas of natural selection and Mendelian genetics were put together with population genetics. The Modern Synthesis also addressed the relationship between the broad-scale changes of macroevolution seen by paleontologists and the small-scale changes of microevolution in local populations of living organisms seen by biologists.
Demography is the statistical study of populations, especially human beings. It encompasses the study of the size, structure, and distribution of these populations, and spatial or temporal changes in them in response to birth, migration, aging, and death.
Darwin, Wallace and natural selection
The actual mechanism for evolution was independently conceived of and described by two naturalists, Charles Darwin and Alfred Russell Wallace (Figure 2.5), in the mid-nineteenth century. Importantly, each spent time exploring the natural world on expeditions to the tropics. From 1831 to 1836, Darwin traveled around the world on H.M.S. Beagle, visiting South America, Australia, and the southern tip of Africa. Wallace traveled to Brazil to collect insects in the Amazon rainforest from 1848 to 1852 and to the Malay Archipelago from 1854 to 1862.
Darwin’s journey, like Wallace’s later journeys in the Malay Archipelago, included stops at several island chains, the last being the Galápagos Islands. On these islands, Darwin observed species on different islands that were clearly similar, yet had distinct differences. For example, the ground finches inhabiting the Galápagos Islands comprised several species that each had a unique beak shape (Figure 2.6). He observed both that these finches closely resembled another finch species on the mainland of South America and that the group of species in the Galápagos formed a graded series of beak sizes and shapes, with very small differences between the most similar. Darwin imagined that the island species might be all species modified from one original mainland species.

Figure 2.5: Charles Darwin in 1854 (left) and Alfred Russell Wallace in 1862 (right)
Figure 2.6: Darwin observed that beak shape varies among finch species. This image shows the beak shape of four species of finch, clockwise (from top left): Geospiza magnirostris (large ground finch) , Geospiza fortis (medium ground finch), Certhidea olivacea (green-warbler finch) , and Camarhynchus parvula (small tree finch) .
Wallace and Darwin both observed similar patterns in other organisms and independently conceived a mechanism to explain how and why such changes could take place. Darwin called this mechanism natural selection. Natural selection, Darwin argued, was an inevitable outcome of three principles that operated in nature:
- Characteristics of organisms are inherited, or passed from parent to offspring.
- More offspring are produced than are able to survive; in other words, resources for survival and reproduction are limited. The capacity for reproduction in all organisms outstrips the availability of resources to support their numbers. Thus, there is a competition for those resources in each generation.
- Offspring vary among each other in regard to their characteristics and those variations are inherited
Both Darwin and Wallace’s understanding of the second principle came from reading an essay by the economist Thomas Malthus, who discussed this principle in relation to human populations. Out of these three principles, Darwin and Wallace reasoned that offspring with inherited characteristics that allow them to best compete for limited resources will survive and have more offspring than those individuals with variations that are less able to compete. Because characteristics are inherited, these traits will be better represented in the next generation. This will lead to change in populations over generations in a process that Darwin called descent with modification.
Papers by Darwin and Wallace presenting the idea of natural selection were read together in 1858 before the Linnaean Society in London. The following year Darwin’s book, On the Origin of Species, was published, which outlined in considerable detail his arguments for evolution by natural selection.
Mendel and inheritance
Johann Gregor Mendel was an Austrian monk in the Augustinian Abbey of St. Thomas in Brno in what is now the Czech Republic. Supported by the monastery, he taught physics, botany, and natural science courses at the secondary and university levels. In 1856, he began a decade of research involving inheritance patterns in pea plants. In 1865, Mendel presented the results of his experiments with nearly 30,000 pea plants to the local natural history society. He demonstrated that traits are transmitted faithfully from parents to offspring in specific patterns. In 1866, he published his work, Experiments in Plant Hybridization, in the proceedings of the Natural History Society of Brünn.
Mendel’s work went virtually unnoticed by the scientific community, which incorrectly believed that the process of inheritance involved a blended inheritance of parental traits that produced an intermediate physical appearance in offspring. This hypothetical process appeared to be correct because of what we know now as continuous variation. Continuous variation is the range of small differences we see among individuals in a characteristic like human height. It does appear that offspring are a “blend” of their parents’ traits when we look at characteristics that exhibit continuous variation. Mendel worked instead with traits that show discontinuous variation. Discontinuous variation is the variation seen among individuals when each individual shows one of two—or a very few—easily distinguishable traits, such as violet or white flowers. Mendel’s choice of these kinds of traits in pea plants allowed him to see experimentally that the traits were not blended in the offspring as would have been expected at the time, but that they were inherited as distinct traits. Mendel was not recognized for his extraordinary scientific contributions during his lifetime; in fact, it was not until 1900 that his work was rediscovered, reproduced, and revitalized by scientists on the brink of discovering the molecular genetic basis of heredity.
Mendel performed hybridizations, which involve mating two true-breeding plants (plants that always produce offspring that look like the parent) with different traits (Figure 2.7). By experimenting with true-breeding pea plants, Mendel avoided the appearance of unexpected traits in offspring that might occur if the plants were not true breeding. Plants used in first-generation crosses were called P, or parental generation, plants. He collected the seeds produced by the P plants that resulted from each cross and grew them the following season. These offspring were called the F1, or the first generation after the parent generation. Once Mendel examined the characteristics in the F1 generation of plants, he allowed them to self-fertilize naturally. He then collected and grew the seeds from the F1 plants to produce the F2, or second generation. Mendel’s
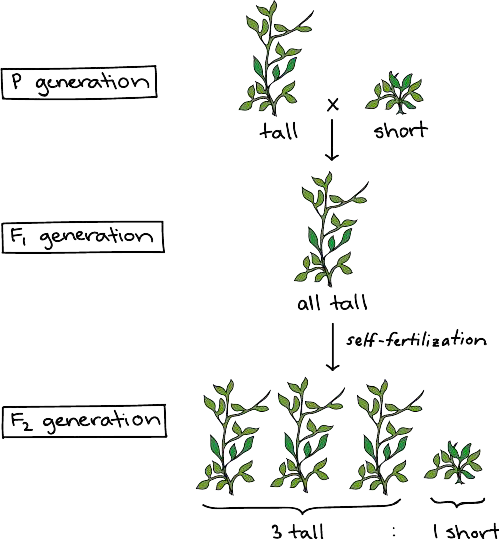
Figure 2.7: Successive generations of pea plants used in Mendel’s experiments
experiments extended beyond the F2 generation to further generations, but it was the 3:1 ratio of characteristics in the P, F1, and F2 generations that were the most intriguing and became the basis of Mendel’s principles of inheritance.
Mendel reported the results of his crosses involving seven different characteristics, each with two contrasting traits. Traits are variations in the physical appearance of heritable characteristics. The characteristics included plant height, seed texture, seed color, flower color, pea-pod size, pea-pod color, and flower position (Figure 2.8). For the characteristic of flower color, for example, the two contrasting traits were white versus purple. To fully examine each characteristic, Mendel generated large numbers of F1 and F2 plants and reported results from thousands of F2 plants.
This 3:1 ratio was no random event. For the other six characteristics that Mendel examined, both the F1 and F2 generations behaved in the same way they did for flower color. One of the two traits would disappear completely from the F1 generation, only to reappear in the F2 generation in a ratio of roughly 3:1.
In Mendel’s experiments, the parent plants pass along discrete units of hereditary information that determine the physical traits of the offspring. We now refer to these units as genes. Each individual has two copies of a given gene, such as the gene for flower color (Figure 2.9). If these copies represent alternate versions, or alleles, of the gene, one allele – the dominant one – may hide the other allele – the recessive one.
The set of alleles carried by an individual is known as its genotype. Genotype and environment determine phenotype, an individual’s observable features. If an individual has two copies of the same allele (say, PP or pp ), it is said to be homozygous for that gene. If, instead, it has two different copies (like Pp ), we can say it is heterozygous.
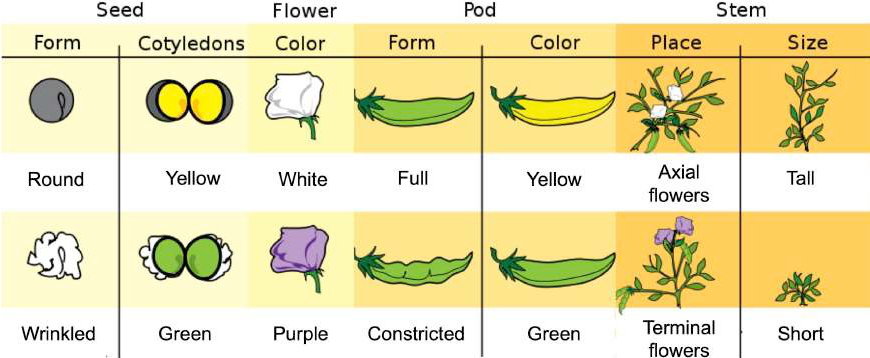
Figure 2.8: Mendel’s seven characteristics and their contrasting traits
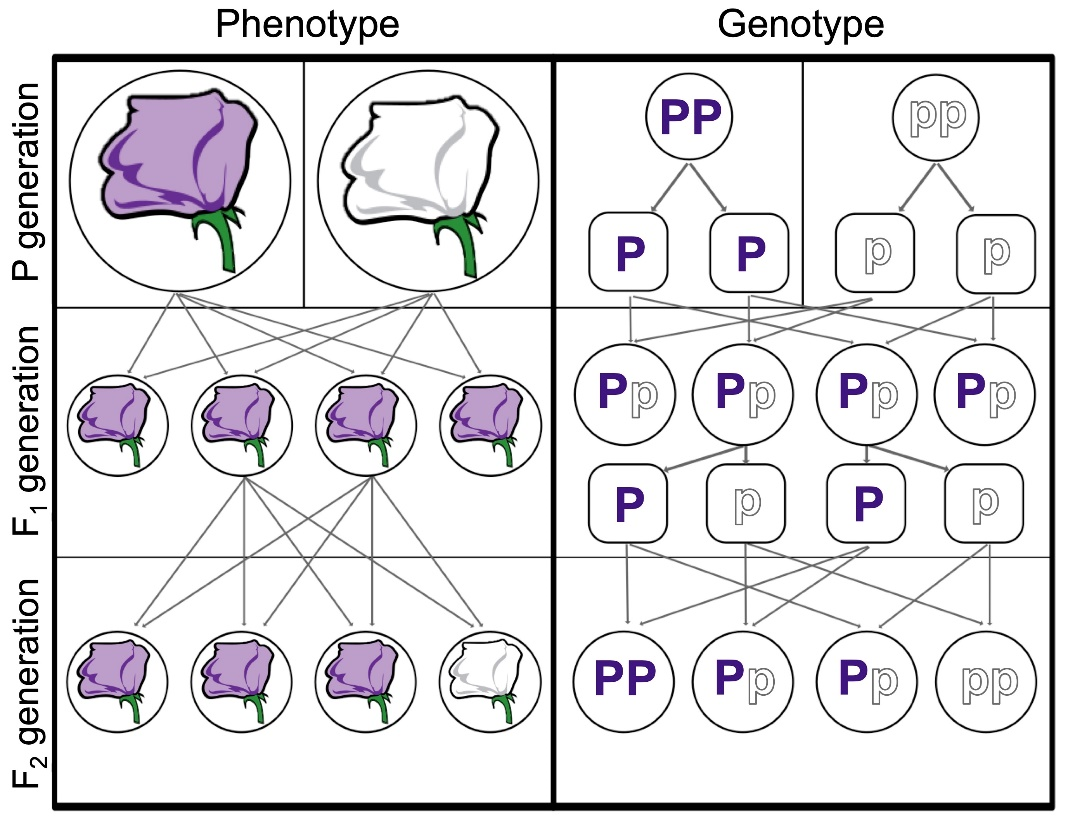
Figure 2.9: For flower color, the dominant allele P hides the recessive allele p. The F1 generation produces 100% purple phenotypes and 100% heterozygous genotypes, while the F2 generation produces 75% purple and 25% white phenotypes (3:1 ratio) and 25% homozygous dominant, 50% heterozygous, and 25% homozygous recessive genotypes.
According to Mendel’s principle of segregation, only one of the two alleles present in an individual is distributed to each gamete (egg or sperm cell) that it makes, and the allocation of the alleles is random. When an egg and a sperm join in fertilization, they form a new individual, whose genotype consists of the alleles contained in the gametes.
The principle of segregation lets us predict how a single feature associated with a single gene is inherited. In some cases, though, we might want to predict the inheritance of two characteristics associated with two different genes. Mendel found that different genes were inherited independently of one another, following what’s called the principle of independent assortment. This principle states that the alleles of two (or more) different genes get sorted into gametes independently of one another. In other words, the allele a gamete receives for one gene does not influence the allele received for another gene.
In Figure 2.10, we can see both the principle of segregation and the principle of assortment illustrated. The allele specifying green pod color is dominant to the allele specifying yellow pod color, and the allele specifying constricted pod form is dominant to the allele specifying full pod form, as shown by the capital and lower-case letters at the bottom of the table. This means that the F1 generation plants would all produce green, constricted pods. Because they are heterozygous for two genes, these F1 generation
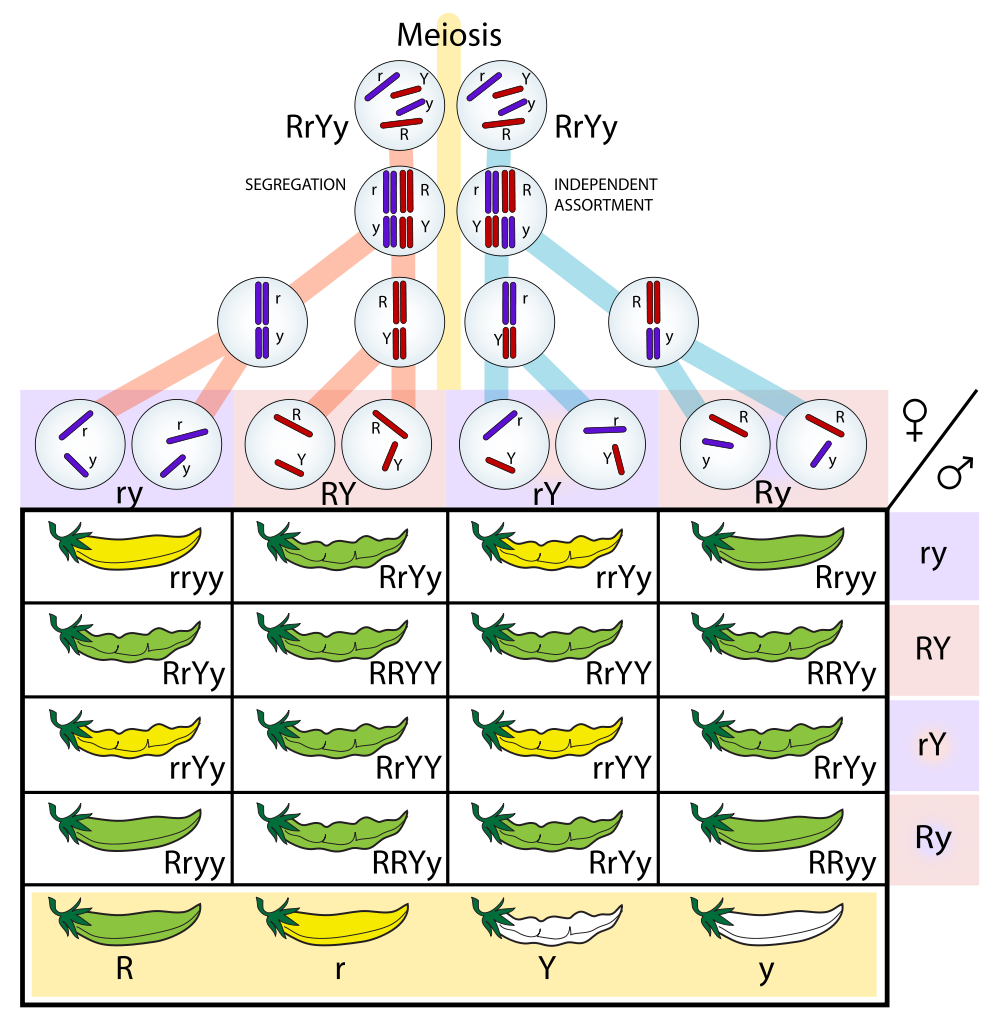
Figure 2.10: Table predicting how two genes ( R and r for pod form, Y and y for pod color) and their alleles separate and then recombine to create variation following Mendel’s principles of segregation and independent assortment during production of gamete cells (meiosis).
plants are called dihybrids. When Mendel experimented with this cross between two dihybrids and looked at the offspring, he found that there were four different categories of pea pods: full and yellow ( rryy ), constricted and yellow ( RRyy ), full and green ( rrYY ), and constricted and green ( RRYY ). These phenotypic categories appeared in a ratio of approximately 9:3:3:1, with no two traits always being passed on together.
Mendel’s principles of segregation and independent assortment and the concepts of dominant and recessive traits are still recognized as key factors in heredity. However, with modern technology we are now able to investigate heredity at the cellular level to determine the inner workings of the units of inheritance.
Molecular Genetics
Heredity is encoded in cells, which are the basic building blocks of all living things. The human body is composed of trillions of cells. They provide structure for the body, but also at the molecular level they take in nutrients from food, convert those nutrients into energy, and carry out specialized functions. Cells can also make copies of themselves.
Cell Structure
All cells fall into two broad categories. Only the single-celled organisms of the domains Bacteria and Archaea are classified as prokaryotes. Animals, plants, fungi, and protists are all multi-celled organisms and are classified as eukaryotes.
Eukaryotic cells have many parts, each with a different function. Some of these parts, called organelles, are specialized structures that perform certain tasks within the cell (Figure 2.11). Human cells contain the following major organelles:
- The nucleus serves as the cell’s command center, sending directions to grow, mature, divide, or die. It also houses DNA (deoxyribonucleic acid), the cell’s hereditary material, and the nucleolus. It is surrounded by the nuclear envelope, which protects the DNA and separates the nucleus from the rest of the cell.
- Ribosomes are organelles that process the cell’s genetic instructions to create proteins. These organelles can float freely in the cytoplasm or be connected to the endoplasmic reticulum.
- Vesicles are a basic tool used by the cell for organizing and transporting cellular substances such as food, wastes and enzymes within the cell.
- Endoplasmic reticulum forms an interconnected network of flattened, membrane-enclosed sacs or tube-like structures. The outer face of rough endoplasmic reticulum is studded with ribosomes that are the sites of protein synthesis. Smooth endoplasmic reticulum lacks ribosomes and functions in lipid synthesis, the production of steroid hormones, and detoxification.
- The Golgi body packages molecules processed by the endoplasmic reticulum to be transported out of the cell.
- The cytoskeleton is a network of long fibers making up the cell’s structural framework. It has several critical functions, including determining cell shape, participating in cell division, and allowing cells to move. It also directs the movement of organelles and other substances within cells.
- Within the cell, cytosol is a gel-like fluid that surrounds organelles.
- Mitochondria are complex organelles that convert energy from food into a form that the cell can use. They have their own genetic material, separate from the DNA in the nucleus, and can make copies of themselves.
- Lysosomes are the recycling center of the cell. They digest foreign bacteria that invade the cell, rid the cell of toxic substances, and recycle worn-out components.
- Centrioles play a key role in determining the position of the nucleus and spatial arrangement of the cell during cell division.
- The cell membrane is the outer lining of the cell. It separates the cell from its environment and allows materials to enter and leave the cell.
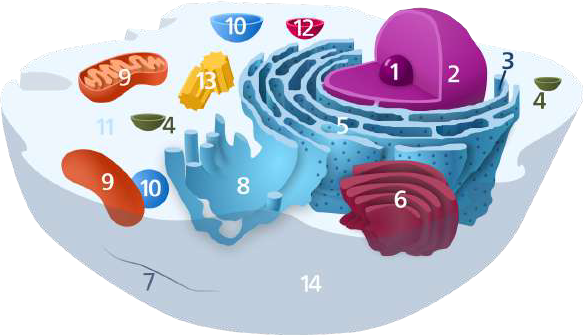
Figure 2.11: Diagram of typical animal cell organelles: 1) nucleolus, 2) nucleus, 3) ribosomes, 4) vesicle, 5) rough endoplasmic reticulum, 6) Golgi body, 7) cytoskeleton, 8) smooth endoplasmic reticulum, 9) mitochondrion, 10) vacuole, 11) cytosol, 12) lysosome, 13) centriole, and 14) cell membrane
Humans and other animals have two kinds of cells: somatic cells, which make up the basic structural components of the body, and gametes, which are sex cells specifically involved in reproduction. Gametes also occur in two forms: ova (female sex cell) and sperm (male sex cell), both of which unite during reproduction to form a zygote with the potential to develop into a new organism.
All multicellular organisms use cell division for growth, and in most cases, the maintenance and repair of cells and tissues. When a cell divides, one of its main jobs is to make sure that each of the two new cells gets a full, perfect copy of genetic material. Mistakes during copying, or unequal division of the genetic material between cells, can lead to cells that are unhealthy or dysfunctional.
Structure of DNA
In the 1950s, Rosalind Franklin, Maurice Wilkins, Francis Crick and James Watson were actively investigating the molecular structure of DNA. Franklin was using X-ray crystallography in Wilkins’ lab to observe the patterns formed by X-rays shot through crystalized DNA fibers. Watson and Crick were able to piece together the puzzle using Franklin’s data and key pieces of information about the pairing of nucleotides in a DNA molecule. In 1953, both Wilkins and Franklin published papers on their X-ray data in the same Nature issue with Watson and Crick’s paper on the structure of DNA. Franklin died from cancer in 1958, and although Franklin’s research on viruses was appreciated in her lifetime, her contributions to the discovery of the structure of DNA were largely recognized posthumously. In 1962, the Nobel Prize in Physiology or Medicine was awarded to Watson, Crick, and Wilkins for solving the structure of DNA.
DNA, or deoxyribonucleic acid, is the hereditary material in humans and almost all other organisms. Nearly every cell in a person’s body has the same DNA. The majority of DNA is located in the cell nucleus (where it is called nuclear DNA ), but a small amount of DNA can also be found in the mitochondria (where it is called mitochondrial DNA or mtDNA). A cell’s complete complement of DNA is called its genome. Normally, DNA molecules are double-stranded, linear, thread-like molecules bound with proteins in the nucleus of each cell. Stretched end-to-end, the DNA molecules in a single human cell are estimated to be about 6 feet in length.
The information in DNA is stored as a code made up of four chemical bases: adenine (A), guanine (G), cytosine (C), and thymine (T). Human DNA consists of about 3 billion bases, and more than 99% of those bases are the same in all people. The order, or sequence, of these bases determines the information available for building and maintaining an organism, similar to the way in which letters of the alphabet appear in a certain order to form words and sentences.
DNA bases pair up with each other, adenine with thymine (A-T) and cytosine with guanine (C-G), to form units called base pairs. Each base is also attached to a sugar molecule and a phosphate molecule. Together, a base, sugar, and phosphate are called a nucleotide. Nucleotides are arranged in two long strands that form a spiral ladder called a double helix. The double helix’s ladder structure is formed with the base pairs as the rungs and the sugar-phosphate molecules as the vertical sidepieces (Figure 2.12).
An important property of DNA is that it serves two functions: to replicate, or make copies of itself, and to produce proteins. During replication (Figure 2.13), each strand of DNA in the double helix can serve as a pattern for duplicating the sequence of bases. This is critical when cells divide because each new cell needs to have an exact copy of the DNA present in the old cell. DNA replication begins with the separation of the bonds holding the base pairs together in the parent strand, creating two template strands.
Figure 2.12: Differing chemical structures of DNA and RNA
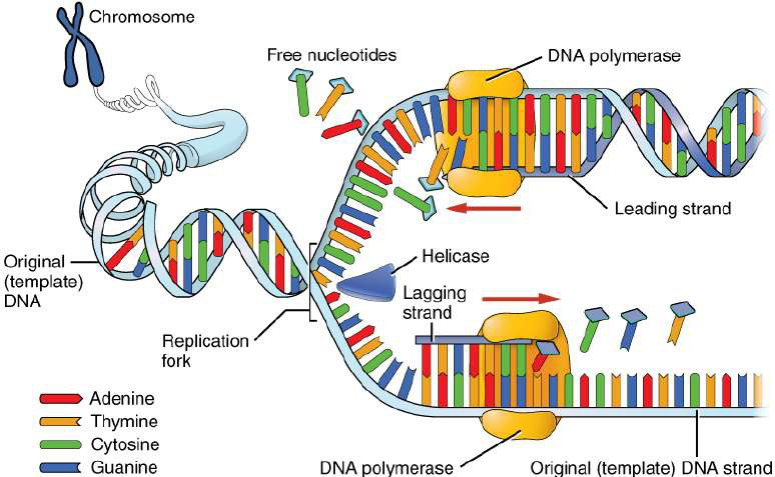
Figure 2.13: Process of DNA replication resulting in two complete DNA molecules
Unpaired bases on the template strands attract and bond with complementary free nucleotides in opposite directions. When all of the new base pairs are formed on each strand, replication is complete, and there are now two complete DNA molecules.
There is a second nucleic acid in all cells called ribonucleic acid, or RNA (Figure 2.12). Like DNA, RNA is made up of nucleotides that include a base, sugar and phosphate. RNA nucleotides contain the chemical bases adenine, cytosine, and guanine, but they do not contain thymine, which is instead replaced by uracil (U). In addition, RNA exists as a single-stranded molecule rather than a double-stranded helix. This molecule plays a key role in transporting amino acids for protein chain construction during protein synthesis.
During the production of proteins, RNA bases are arranged in sequences of three, which are called codons. These sequences determine the assembly of different amino acids. For example, the combination and arrangement of thymine, adenine, and guanine encode the amino acid isoleucine, which helps rebuild muscle tissue. Amino acids join together in chains to produce different proteins, which are chemical compounds that are necessary for the structure and operational processes of the body’s cells. There are 20 different amino acids that can come together in differing combinations and amounts to produce millions of different proteins basic to life.
Chromosomes and cell division
Gametes and somatic cells divide to produce new cells in different ways. While both kinds of cells contain an individual’s genome, only the gametes provide the means for passing this information on to the next generation. During cell division, the DNA molecule becomes tightly coiled into structures known as chromosomes. In humans, somatic cells are diploid, having a full complement of 46 chromosomes (23 matched pairs), while gametes are haploid with 23 chromosomes, or half the number of a diploid cell.
The matched pairs of chromosomes in a diploid cell are called homologous chromosomes. Homologous chromosomes are the same length and have genes in the same locations. The sex chromosomes, X and Y, determine a person’s biological sex: XX specifies female and XY specifies male. These chromosomes are not truly homologous and are an exception to the rule of the same genes in the same places. Aside from small regions of similarity needed during sex cell production, the X and Y chromosomes are different and carry different genes. The 44 non-sex chromosomes in humans are called autosomes.
Somatic cells are produced through the process of mitosis (Figure 2.14, top). Mitosis is divided into a series of phases that result in the division of the cell nucleus and two new diploid cells with 46 chromosomes. In contrast, gamete cells are produced through the process of meiosis (Figure 2.14, bottom), or two successive cell divisions that result in four new haploid cells with 23 chromosomes. Meiosis reduces the number of chromosomes to prepare for sexual reproduction, thus, when a zygote is formed from
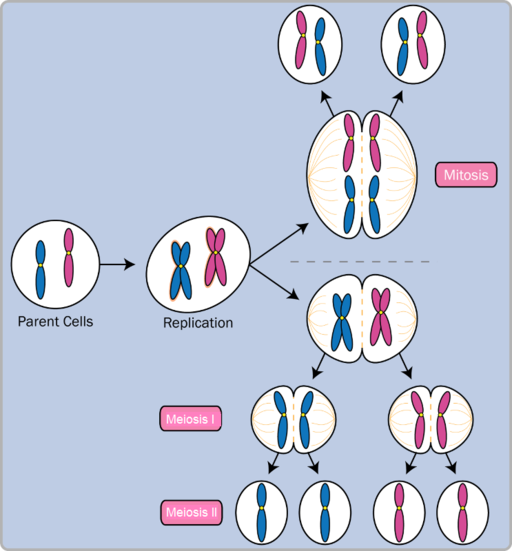
Figure 2.14: Comparison of the processes of mitosis (top) and meiosis (bottom) and their resulting new haploid and diploid cells after cell division.
male and female gametes, a full complement of 46 chromosomes are present to produce a new individual. Nondisjunction events occur randomly when the sex chromosomes fail to separate normally during meiosis, producing a sperm with an extra X or Y chromosome or an ovum with two X chromosomes. Nondisjunction can also result in a missing or partly altered sex chromosome. In humans, common survivable nondisjunction events include:
- Klinefelter’s syndrome, where 1:1000 males has an extra X chromosome (XXY)
- Jacobs syndrome, where 1:1000 males has an extra Y chromosome (XYY)
- Trisomy X, where 1:1000 females has an extra X chromosome (XXX)
- Turner syndrome, where 1:2000 females are partly or completely missing an X chromosome (X0)
Unlike Mendel’s tall or short plants and green or yellow peas, humans don’t necessarily have such distinct phenotypic expressions of traits. These kinds of inherited traits are referred to as traits of simple inheritance. Humans share approximately 4,500 traits that are inherited in this manner, but the majority of our traits are passed on in a more complicated way. Height and other similar features are controlled not just by one gene, but rather, by multiple (often many) genes that contribute in some way to the overall outcome. This inheritance pattern is called polygenic inheritance. These kinds of traits form a gradation and are influenced by alleles of multiple genes. They include many of our most prominent phenotypic traits, such as hair color, eye color, skin color, which all produce a wide spectrum of variants.
Genetic traits and their expression are also influenced by many other factors that are external to the DNA sequence. In some cases, the genetic instructions provided by genes may be turned “on” or “off” and subsequently affecting whether the cells should produce relevant proteins. For example, muscle and nerve cells contain the same DNA, but appear different because their genes are modified in varied ways. Epigenetics is the study of changes in organisms caused by modification of gene expression rather than alteration of the genetic code itself. The different combinations of genes that are turned on or off is what makes each one of us unique. Furthermore, there have been indications that some epigenetic changes can be inherited.
Population Genetics and Evolution
Recall that a gene for a particular character may have several variants, or alleles, that code for different traits associated with that character. Each individual in a population of diploid organisms can only carry two alleles for a particular gene, but more than two may be present in the individuals that make up the population. In the early twentieth century, biologists began to examine what happens to all the alleles in a population in a field of study known as population genetics.
Until now, we have defined evolution as a change in the heritable characteristics of biological populations over successive generations, but behind that phenotypic change is genetic change. In population genetics terms, microevolution involves how allele frequencies change over time in a population.
There are several ways the allele frequencies of a population can change. A heritable trait that aids the survival and reproduction of an organism in its present environment is called an adaptation. Adaptation to an environment comes about when a change in the range of genetic variation occurs over time that increases or maintains the match of the population with its environment. Whether or not a trait is favorable depends on the environment at the time. The same traits do not always have the same relative benefit or disadvantage because environmental conditions can change.
If a given allele confers a phenotype that allows an individual to have more offspring that survive and reproduce, that allele, by virtue of being inherited by those offspring, will be in greater frequency in the next generation. This kind of reproductive success is referred to as evolutionary fitness. Since allele frequencies in a population always add up to 100%, an increase in the frequency of one allele always means a corresponding decrease in one or more of the other alleles. Highly beneficial alleles may, over a very few generations, become “fixed” in this way, meaning that every individual of the population will carry the allele. Similarly, detrimental alleles may be swiftly eliminated from the gene pool, the sum of all the alleles in a population.
Selective pressure is any external force that causes differences in the fitness of individuals having particular alleles. A part of the study of population genetics is tracking how selective pressures change the allele frequencies in a population over time, which can give scientists clues regarding the mechanisms that may be operating on a given population.
Mechanisms of change
In the early twentieth century, English mathematician Godfrey Hardy and German physician Wilhelm Weinberg independently provided an explanation for a somewhat counterintuitive concept. Hardy’s original explanation was in response to a misunderstanding as to why a “dominant” allele, one that masks a recessive allele, should not increase in frequency in a population until it eliminated all the other alleles. The question resulted from a common confusion about what “dominant” means, but it forced Hardy to mathematically point out that if there are no evolutionary mechanisms acting on the population and affecting allele frequencies, those frequencies in the population will remain constant in the same proportions from one generation to the next. This principle is now known as the Hardy-Weinberg equilibrium. If this were the case, individuals would, as a whole, look essentially the same, and this would be unrelated to whether the alleles were dominant or recessive.
The evolutionary mechanisms of change acting on populations are natural selection, mutation, genetic drift, and gene flow (migration). In fact, we know they are probably always affecting populations, independent of each other or in combination. Natural selection is the process that produces adaptations and exerts pressures that favor some individual over others to survive and reproduce in a given environment (Figure 2.15). An individual is born with the genes it has – these do not change as the individual ages. Therefore, an individual cannot evolve or adapt through natural selection. Natural selection can only take place if there is variation, or differences among individuals in a population. Importantly, these differences must have some genetic basis; otherwise, selection will not lead to change in the next generation. This is critical because variation among individuals can be caused by non-genetic reasons, such as an individual being taller because of better nutrition rather than different genes.
Mutation, a change in the DNA sequence of a gene, is the ultimate source of new alleles or new genetic variation in any population. Mutations can occur spontaneously during DNA replication, especially during meiosis, or they can be induced through other types of
damage to DNA, such as exposure to carcinogens or radiation. A mutation can have one of three outcomes:
- A mutation may affect the phenotype of an organism in a way that reduces fitness.
- A mutation may produce a phenotype with a beneficial effect on fitness.
- Many mutations, called neutral mutations, will have no effect on fitness.
A mutation can change an allele, but the net effect is a change in frequency. The change in frequency resulting from mutation is small, so its effect on evolution is small unless it interacts with one of the other factors, such as selection (Figure 2.16). Harmful mutations
Figure 2.15: A population of mice undergoing the process of natural selection
Figure 2.16: A white peacock with a genetic mutation known as leucism, which causes the lack of pigments in the plumage; this may affect the individual’s selection as a possible mate within the population
are removed from the population by selection and will generally only be found in very low frequencies equal to the mutation rate. Beneficial mutations will spread through the population through selection, although that initial spread is slow. Whether or not a mutation is beneficial or harmful is determined by whether it helps an organism survive to sexual maturity and reproduce.
Random differences in the frequency of an allele within a small or isolated population due to chance events are called genetic drift. Genetic drift does not introduce new alleles to a population, but it can reduce variation within a population by removing an allele from the gene pool (Figure 2.17). This outcome occurs because the alleles in an offspring generation are a random sample of the alleles in the parent generation. Alleles may or may not make it into the next generation due to chance events including mortality of an individual, events affecting finding a mate, and even the events affecting which gametes end up in fertilizations.
Genetic drift has a much larger impact on smaller populations than larger populations. Imagine a population with 10 individuals. If one individual in the population happens to die before it leaves any offspring to the next generation, all of its genes—10% of the population’s gene pool—will be suddenly lost. In a population of 100, that one
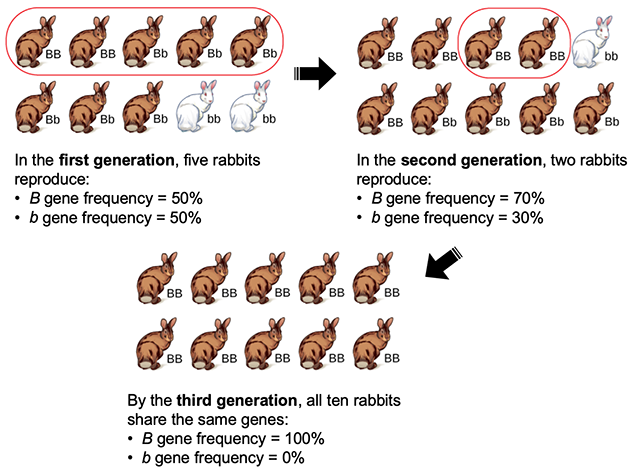
Figure 2.17: Genetic drift can lead to elimination of an allele by chance; in each generation, a random set of individuals produce the next generation, and the frequency of alleles in the generation is equal to the frequency of alleles among those reproducing
individual represents only 1% of the overall gene pool; therefore, it has much less impact on the population’s genetic structure and is unlikely to remove all copies of even a relatively rare allele.
Genetic drift can also be magnified by natural or human-caused events. In one fell swoop, the genetic structure of the survivors becomes the genetic structure of the entire population, which may be very different from the pre-disaster population. This type of event is known as a population bottleneck (Figure 2.18).
Bottleneck effect occurs when the population of a species is reduced drastically over a short period of time due to external forces. In a true population bottleneck, the reduction does not favor any combination of alleles; it is totally random chance which individuals survive. In other words, the disaster must be one that kills for reasons unrelated to the organism’s traits, such as a lava flow or hunting an animal almost to extinction. A bottleneck can reduce or eliminate genetic variation from a population. Further genetic drift events after the bottleneck event can also reduce the population to having little to no genetic diversity. The lack of diversity created can make the population at risk to other selective pressures.
Populations might also experience a strong influence or magnification of genetic drift. In cases where a part of the population leaves to start a new population in a new location, or if a population is divided by some kind of physical barrier, and each portion of the
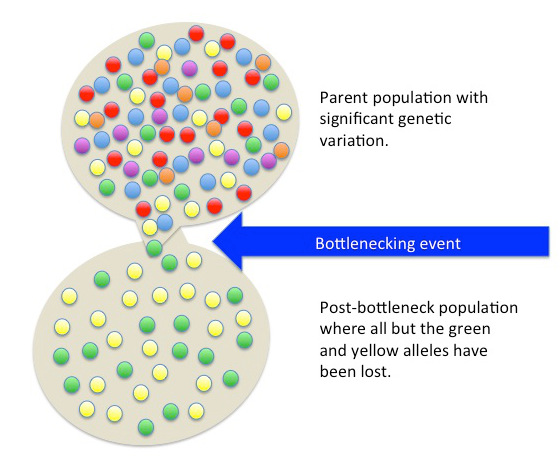
Figure 2.18: A chance bottleneck event can reduce the genetic variation within a population through the loss of alleles.
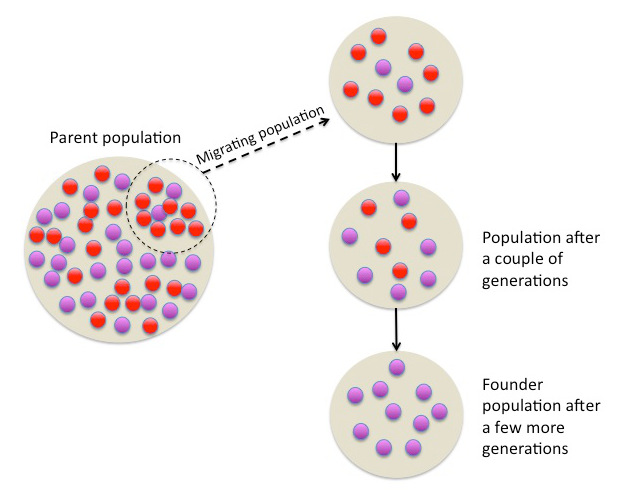
Figure 2.19: Founder effect resulting from the changing frequency of a random gene over generations in a small population isolated from the parent population.
original population becomes isolated, individuals in the new population are unlikely to be representative of the ‘parent’ population. Over generations, this results in a founder effect (Figure 2.19). Founders of the new population will determine the genetic makeup, and potentially the survival, of the population for generations. For example, the Afrikaner population of Dutch settlers in South Africa is descended mainly from a few colonists. Today, the Afrikaner population has an unusually high frequency of the gene that causes Huntington’s disease, because those original Dutch colonists just happened to carry that gene with unusually high frequency. This effect is easy to recognize in genetic diseases, but of course, the frequencies of all sorts of genes are affected by founder events.
The final important mechanism of change is gene flow (Figure 2.20), or the exchange of genes between interbreeding populations. Gene flow affects populations in some very important ways. The introduction of new genes into a population changes the allele frequencies in that population, increasing genetic variation. However, gene flow can also decrease variation between populations. If two distant populations continue to interbreed, they will become genetically similar to one another. Human populations have experienced extensive gene flow for millennia, preventing substantial genetic distance between populations, which helps to explain why there is only one human species today.
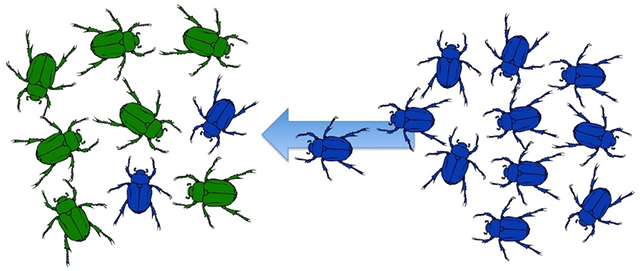
Figure 2.20: Gene flow occurs when individuals with a gene ( e.g., blue color) migrate from one location to another and interbreed with a different population ( e.g., green color) of the same species.
Process of speciation
In biology and biological anthropology, a species is often defined as the largest taxonomic group of organisms where any two individuals of the appropriate sexes or mating types can produce fertile offspring, typically by sexual reproduction. There are exceptions to this rule. Many species are similar enough that hybridization (Figure 2.21), or the process of reproducing offspring by combining the qualities of two organisms of different species, is possible and often occurs in nature, but for the majority of species this rule generally holds. In fact, the presence of hybrids between similar species suggests that they may have descended from a single interbreeding species and that the process of speciation may not yet be completed. In many instances hybrid offspring are not reproductively fit; however, if the parent species produce a hybrid that is reproductively successful, the hybrid population may not be able to reproduce with the parent species, resulting in a speciation event.
Figure 2.21: Process of hybridization where two species produce evolutionarily fit hybrid offspring that are unable to reproduce with members of its parent species.
For this reason, other concepts in defining species include their number and appearance of chromosomes, DNA sequence, morphology, behavior or ecological niche, and make no claim about reproductive isolation, making the concept useful where only fossil evidence is available.
To examine the evolutionary relationships among species, evolutionary biologists construct phylogenies. Many species have similar traits because they are descendants of a single common ancestor. These species developed from a single source and are related to a certain degree despite their current differences. The traits they share are known as homologous structures. Homologous structures are similar in structure and function because they originated from the same ancestor long ago. Examples of homologous structures (Figure 2.22) are the bones in the forelimbs of various vertebrates, such as humans, cats, seals and bats. Even though the functions of these limbs are different (a human hand is for grasping, a cat’s foot is for walking, a seal’s flipper is for swimming, and a bat’s wing is for flying), they all have similar bone structures.
Species may also have similar morphological traits even though they are not related to each other. This usually results because the species live in similar environments and fill similar ecological roles. The structures in this case are known as analogous structures. Examples of analogous structures range from wings in flying animals like bats, birds, and insects, to fins in animals like penguins and fish.
Figure 2.22: Homologous bones in the structures of vertebrates; all of these animals have the same basic structure, but have been adapted for specific uses.
Evolutionary changes above the species level are referred to as macroevolution. Two types of macroevolution are divergent evolution and convergent evolution. Divergent evolution occurs when two separate species evolve differently from a common ancestor. An example of divergent evolution can be seen in New World and Old World monkeys, which share basic anatomies; however, they can look very different as a result of geographic isolation, and adapting to different environments.
Convergent evolution occurs when species have different ancestral origins but have developed similar features. It is important to understand convergent evolution; otherwise, we might mistakenly assume a relationship between two animals that have similar features. For example, hummingbirds and hummingbird moths are the same size, eat nectar from the same flowers, and have wings that work in a similar manner. However, one is an insect and the other is a bird, and they did not evolve from a common ancestor. Instead, each evolved from different ancestors in a similar environment and developed the same characteristics.
Each time one ancestral species diverges into multiple descendant species it is called speciation. For this process to occur, two new populations must be formed from one original population, and they must evolve in such a way that it becomes impossible for individuals from the two new populations to interbreed. There are two broad modes of how this could occur. The first is allopatric speciation (or vicariance ), which occurs when biological populations of the same species become isolated from each other to an extent that prevents or interferes with gene flow (Figure 2.23). An example of allopatric speciation is seen in ring species, where adjacent populations of the same species are able to reproduce around a barrier, but gradual changes in the intermediate populations in the series result in two different species no longer able to successfully interbreed (Figure 2.24).
Figure 2.23: Process of allopatric speciation where a species population becomes separated by a geographic barrier and the reproductive isolation results in two separate species
Figure 2.24: Process of ring speciation, where adjacent populations of the same species are able to reproduce around a barrier, but gradual changes in intermediate populations result in two different species unable to successfully interbreed
Figure 2.25: Process of sympatric speciation where reproductive isolation within a population occurs in the same geographic range without geographic barriers
The second broad mode of speciation is sympatric speciation, which is the isolation and evolution of a new species from a surviving ancestral species while both species continue to inhabit the same geographic region (Figure 2.25). An example of sympatric speciation can be seen in apple flies. Hawthorn apple flies used to lay their eggs only on the fruit of hawthorn trees (native to North America), but less than 200 years ago, some hawthorn flies began to lay their eggs on domestic apples (introduced to North America by migrants). Now there are two groups of apple flies: one that lays eggs on hawthorns and one that lays eggs on apples. Males look for mates on the same type of fruit that they matured on, and females lay their eggs on the same type of fruit that they matured on. Therefore, flies that matured on hawthorns will raise offspring on hawthorns, and flies that matured on apples will raise offspring on apples. There are genetic differences between the two groups, and over a long period of time, they are likely to become separate species.
In some cases, a population of one species disperses throughout an area, and each finds a distinct niche or isolated habitat. Over time, the varied demands of their new lifestyles lead to diversification and ultimately multiple speciation events originating from a single species, or adaptive radiation. From one point of origin, many adaptations evolve causing the species to radiate into several new ones. An excellent example of this is seen in the diversity of beak shapes of Darwin’s finches (review Figure 2.6). Each finch beak is adapted for a specialized feeding behavior.
Biotechnology and Genetics
The latter half of the twentieth century began with the discovery of the structure of DNA, then progressed to the development of the basic tools used to study and manipulate DNA. These advances, as well as advances in our understanding of and ability to manipulate cells, have led some to refer to the twenty-first century as the biotechnology century. The rate of discovery and of the development of new applications in medicine, agriculture, and energy is expected to accelerate, bringing huge benefits to humankind and perhaps also significant risks. Many of these developments are expected to raise significant ethical and social questions that human societies have not yet had to consider.
Biotechnology is the use of artificial methods to modify the genetic material of living organisms or cells to produce novel compounds or to perform new functions. Since the discovery of the structure of DNA in 1953, and particularly since the development of tools and methods to manipulate DNA in the 1970s, biotechnology has become synonymous with the manipulation of organisms’ DNA at the molecular level.
Cloning means the creation of a perfect replica. Typically, the word is used to describe the creation of a genetically identical copy. Reproductive cloning is a method used to make a clone or an identical copy of an entire multicellular organism. Most multicellular organisms undergo reproduction by sexual means, which involves the contribution of DNA from two individuals (parents), making it impossible to generate an identical copy or a clone of either parent. Long before attempts were made to clone an entire organism, researchers learned how to copy short stretches of DNA—a process that is referred to as molecular cloning. Molecular cloning allows for the creation of multiple copies of genes, expression of genes, and study of specific genes.
The classic methods of studying the function of genes began with a given phenotype and determined the genetic basis of that phenotype, but modern techniques allow researchers to start at the DNA sequence level and ask: “What does this gene or DNA element do?” This technique, called reverse genetics, has resulted in reversing the classical genetic methodology. In a reverse genetics approach, mutating or deleting genes provides researchers with clues about gene function. Alternately, reverse genetics can be used to cause a gene to overexpress itself to determine what phenotypic effects may occur.
Medicine and genetics
It is easy to see how biotechnology and genomics can be used for medicinal purposes. Knowledge of the genetic makeup of our species, the genetic basis of heritable diseases, and the invention of technology to manipulate and fix mutant genes provides methods to treat diseases. The process of testing for suspected genetic defects before administering medical treatment is called genetic diagnosis by genetic testing. In cases in which a genetic disease is present in an individual’s family, family members may be advised to undergo genetic testing. Gene therapy is an experimental genetic engineering technique that may one day be used to treat, cure or prevent certain genetic diseases. In its simplest form, it involves the introduction of a non-mutated gene at a random location in the genome to cure a disease by replacing a protein that may be absent in these individuals because of a genetic mutation. To date, gene therapies have been primarily experimental procedures in humans. A few of these experimental treatments have been successful, but the methods may be important in the future as the factors limiting its success are resolved.
Although there have been significant advances in the medical sciences in recent years, doctors are still confounded by many diseases and researchers are using whole genome sequencing to get to the bottom of the problem. Whole genome sequencing is a process that determines the DNA sequence of an entire genome. Whole genome sequencing is a brute-force approach to problem solving when there is a genetic basis at the core of a disease. Several laboratories now provide services to sequence, analyze, and interpret entire genomes.
DNA sequencing technology has led to great advances in genomics. Just as information technology has led to Google Maps enabling us to get detailed information about locations around the globe, genomic information is used to create similar maps of the DNA of different organisms. Genome mapping is the process of finding the location of genes on each chromosome. The maps that are created are comparable to the maps that we use to navigate streets. A genetic map (Figure 2.26) is an illustration that lists genes and their location on a chromosome. Genetic maps provide the big picture (similar to a map of interstate highways) and use genetic markers (similar to landmarks). A genetic marker is a gene or gene sequence on a chromosome that shows genetic linkage with a trait of interest. The genetic marker tends to be inherited with the gene of interest, and one measure of distance between them is the recombination frequency during meiosis. Early geneticists called this linkage analysis.
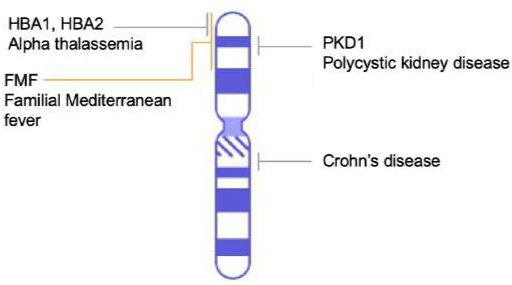
Figure 2.26: Genetic map of human chromosome 16 listing location of genes associated with specific diseases.
Having a complete map of the genome makes it easier for researchers to study individual genes. Human genome maps help researchers in their efforts to identify human disease-causing genes related to illnesses such as cancer, heart disease, and cystic fibrosis, to name a few. In addition, genome mapping can be used to help identify organisms with beneficial traits, such as microbes with the ability to clean up pollutants or even prevent pollution.
Genomic mapping is used with different model organisms that are used for research. This is still an ongoing process, and, as more advanced techniques develop, more advances are expected. Genome mapping is similar to completing a complicated puzzle using every piece of available data. Mapping information generated in laboratories all over the world is entered into central databases, such as the National Center for Biotechnology Information (NCBI). Efforts are made to make the information more easily accessible to researchers and the general public. Just as we use global positioning systems instead of paper maps to navigate through roadways, NCBI allows us to use a genome viewer tool to simplify the data mining process.
Science, Culture and Evolution
Science is the study of the natural world based on facts, or things that are proved to be true through experiments and observation. In contrast, cultural constructions are the cognitive structures people create in order to perceive and classify aspects of the natural and social world around them. Cultural constructions are not based on facts, but instead on personal and group belief, or trust in the existence of something not susceptible to rigorous proof. Evolutionary theory is science, while belief is a cultural construction.
Critics of the science of evolution attempt to dismiss its importance by purposefully confounding everyday usage of the word ‘theory’ with the way scientists use the word. In science, a theory is understood to be a concept that has been extensively tested and supported over time. We have a theory of the atom, a theory of gravity, and the theory of relativity, each of which describes what scientists understand to be facts about the world. In the same way, the theory of evolution describes facts about the natural world. As such, a theory in science has to survive significant efforts to discredit it by scientists, who are naturally skeptical. While theories can sometimes be overturned or revised, this does not lessen their weight but rather simply reflects the constantly evolving state of scientific knowledge. In contrast, a ‘theory’ in common everyday usage means a guess or suggested explanation for something. This everyday meaning is more similar to the concept of a hypothesis, which, in science, is a proposed explanation for a phenomenon to be supported or disproved. When critics of the science of evolution say evolution is “just a theory,” they are implying that there is little evidence supporting it and mischaracterizing it as still in the process of being rigorously tested. Therefore, evolution is an accepted and settled science.
There are two common misconceptions concerning the scientific theory of evolution. The first is that evolution includes an explanation of life’s origins. The science in this theory does not try to explain the origin of life, but instead explains how populations change over time and how life diversifies. The second misconception is situated in statements such as “organisms evolve in response to a change in an environment,” which are quite common. This should not be understood to mean that individual organisms evolve, as discussed earlier in this chapter. The statement is shorthand for “a population evolves in response to a changing environment.” However, this can also lead to additional misunderstanding by interpreting the statement to mean that evolution is somehow intentional. A changed environment results in some individuals in the population, those with particular phenotypes, benefiting and, therefore, producing proportionately more offspring than other phenotypes. This results in change in the population only if the characteristics are genetically determined.
In a larger sense, evolution is also not goal directed. Species do not become “better” over time; organisms simply adjust to their changing environment with adaptations that maximize their reproduction in a particular environment at a particular time. Evolution has no goal of making faster, bigger, more complex, or even smarter species. This kind of language is common in popular literature. Certain organisms, humans included, are described as the “pinnacle” of evolution, or “perfected” by evolution. What characteristics evolve in a species are a function of the variation present and the environment, both of which are constantly changing in a non-directional way. A trait that is fit in one environment at one time may well be fatal at some point in the future. This holds equally well for a species of insect as it does for the human species.
Evolution and education
After 1918 in the aftermath of World War I, American society experienced a rise of evangelicalism and wide support for certain conservative and revitalization movements in part due to increased use of public education systems, distrust of government war actions and newly formed internal policing agencies, and economic pressures of the Great Depression. These movements and social concerns included a surge of opposition to the scientific evidence supporting the theory of evolution, with several regions of the United States introducing legislation prohibiting the teaching of evolution. By 1925, such legislation was being considered in 15 states, and had passed in some states, such as Tennessee. The American Civil Liberties Union offered to defend anyone who wanted to bring a test case against one of these laws. In July 1925, John T. Scopes (Figure 2.27), a substitute high school teacher, accepted and confessed to teaching his Tennessee science class evolution in defiance of Tennessee’s Butler Act, which had made it unlawful to teach human evolution in any state-funded school. The State of Tennessee v. John Thomas Scopes is commonly referred to as the Scopes Trial.
The trial served its purpose as a publicity stunt, but also as a way to amplify divisions and create similar opportunities in other locations. It drew intense national publicity as reporters flocked to the small town of Dayton to cover the big-name lawyers representing each side. William Jennings Bryan, a three-time presidential candidate, argued for the prosecution, while Clarence Darrow, a famed defense attorney, spoke for Scopes. The widespread publicity of the trial galvanized proponents of evolution by

Figure 2.27: John Thomas Scopes prior to his trial in 1925.
highlighting the controversy between Modernists, who argued that the theory of evolution was not inconsistent with religion, and Fundamentalists, who argued that the word of God as revealed in the Bible took priority over all human knowledge. The trial revealed a growing chasm in American society emphasizing and escalating the political, cultural and legal conflicts between approaches to finding the “truth.” This heavily influenced the extent to which the science of evolution would be taught in the United States.
Creationism is the religious belief that nature, and aspects such as the universe, Earth, life, and humans, originated with supernatural acts of divine creation. In its broadest sense, creationism includes a continuum of religious views, which vary in their acceptance or rejection of scientific explanations, such as evolution, that describe the origin and development of natural phenomena. After the Scopes Trial, various creationism groups in other states sought to pass anti-evolution laws similar to Tennessee’s. By 1927, there were 13 states, both in the North and South, considering some form of anti-evolution law. Nearly all of these efforts were rejected, but Mississippi and Arkansas did put anti-evolution laws on the books that would outlive the Tennessee Butler Act.
Through the following decades of the 20th century, many American anti-evolution crusaders sought to ban evolution as a science topic for study in public schools or, failing that, to relegate it to the status of unproven hypothesis perhaps taught alongside a biblical version of creation, while the majority of educators, scientists, and other professional laymen favored the theory of evolution. In 1958 the National Defense Education Act was passed with the encouragement of many legislators who feared that science and mathematics in the United States education system was falling behind that of the Soviet Union. The act yielded textbooks, produced in cooperation with the American Institute of Biological Sciences, which stressed the importance of evolution as the unifying theory of biology.
Opponents to the theory of evolution made a transition from the creationism anti-evolution crusade of the 1920s to a creation “science” movement in the 1960s, which was a pseudoscientific attempt to use the Bible as scientific fact and nullify scientific evidence for evolution. Despite similarities between these two causes, creation “science” was a shift from overtly religious to covertly religious objections to evolutionary theory. In the United States, texts and curricula that emerged from this later movement focused on concepts derived from a literal interpretation of the Bible, most notably linking the flood in the Biblical Genesis account to the geological and fossil record. These works attracted little notice beyond the American schools and congregations of fundamental Christianity and social conservatives until the 1970s, when its followers challenged the teaching of the theory of evolution in public schools and other science venues in the United States, bringing it to the attention of the public-at-large and the scientific community.
Various state laws prohibiting teaching of evolution were overturned in 1968 when the United States Supreme Court ruled in Epperson v. Arkansas that such laws violated the First Amendment to the United States Constitution. This ruling inspired a new creationist movement to promote laws requiring that schools give balanced treatment to scientific creationism when evolution is taught. The 1981 Arkansas Act 590 was one such law that carefully detailed the principles of scientific creationism that were to receive equal time in public schools alongside evolutionary principles.
The overwhelming consensus of the scientific community is that creation “science” is based on beliefs mixed with misinterpretations of scientific fact. It fails to qualify as a science because it lacks empirical support, supplies no tentative hypotheses or facts, and resolves to describe natural history in terms of scientifically untestable supernatural causes. Courts have also ruled that creation “science” is a religious view base on beliefs rather than scientific fact. The 1982 ruling in McLean v. Arkansas found that creation “science” fails to meet the essential characteristics of science. The teaching of creation “science” in public schools in the United States effectively ended in 1987 following the United States Supreme Court decision in Edwards v. Aguillard, affirming that a Louisiana statute requiring the teaching of creation “science’ alongside evolution when evolution is taught in public schools was unconstitutional because its sole true purpose was to advance a particular religious belief.
Another form of creation “science,” referred to as intelligent design, argues that the complexity of life cannot be explained by modern science and that certain biological features are too complex to be the result of natural processes; therefore, wherever complex design exists, there must have been a designer, and since nature is complex, nature must have had an intelligent designer. Central to the argument posed by proponents of intelligent design is the concept of irreducible complexity, or the pseudoscientific idea that certain biological systems cannot evolve by successive small modifications to pre-existing functional systems through natural selection. This concept is based on the mistaken assumption that evolution relies on improvement of existing functions, ignoring how complex adaptations originate from changes in function, and disregards established scientific facts supporting the theory of evolution. In 2005, requiring intelligent design to be taught in public school science classes was found to be unconstitutional in the Kitzmiller v. Dover Area School District federal court case.
While controversy concerning the teaching of evolution in schools is still prominent in American social, cultural and political arenas, creationism is more widely rejected in Canada, Australia, and Europe. However, in the early- and mid-2000s, there continued to be scattered and increased efforts on the part of religious groups throughout Europe to introduce creationism into public education. In response, the Parliamentary Assembly of the Council of Europe released a draft report titled The Dangers of Creationism in Education in June 2007, which was reinforced by a further proposal of banning it in schools later than same year.
Review of Learning Objectives
Evolution by natural selection arises from three conditions: individuals within a species vary, some of those variations are heritable, and organisms have more offspring than resources can support. The evidence for evolution is found at all levels of organization in living things and in known extinct species. Anatomy of species reveals common structures in divergent lineages that have been modified over time by evolution. The geographical distribution of living species reflects the origins of species in particular geographic locations and the history of movements. The structures of molecules, like anatomical structures, reflect the relationships of living species and match patterns of similarity expected from descent with modification.
The Modern Synthesis of evolutionary theory grew out of the reconciliation of earlier scientific thoughts on evolution and heredity. Population genetics is a theoretical framework for describing evolutionary change in populations through the change in allele frequencies. Population genetics defines evolution as a change in allele frequency over generations. In the absence of evolutionary forces allele frequencies will not change in a population; however, in all populations, mutation, natural selection, genetic drift, and migration act to change allele frequencies.
Genes are inherited as pairs of alleles that behave in a dominant and recessive pattern. Alleles segregate into gametes with each gamete equally likely to receive either one of the two alleles present in a diploid individual. In addition, genes are distributed into gametes independently of one another. The technology of molecular genetics developed in the last half century with the scientific understanding of DNA. This development has enabled us to see deep into the history of life to deduce the relationships between living things in ways never thought possible. It also allows us to understand the workings of evolution in populations of organisms. Thousands of species, including humans, have had their entire genome sequenced. These sequences will allow us to understand human disease and the relationship of humans to the rest of life on Earth.
The factual and scientific nature of evolution is often challenged by wrongly associating the scientific meaning of “theory” with the everyday usage and meaning of the term. Evolution is mistakenly interpreted to mean that individuals evolve, when in fact only populations can evolve as their gene frequencies change over time. Evolution does not explain the origin of life, nor is it goal-directed with organisms changing through intention; however, characteristics that evolve in species are widely recognized and accepted as a function of the variation present in organisms and the environment, both of which are constantly changing in an adaptive, non-directional process.
Concept review
- List six scientific disciplines that played key roles in the development of the theory of evolution by natural selection.
- Describe the three main concepts that grew into the Modern Synthesis of evolutionary theory.
- Describe the structures that are characteristic of a eukaryote cell.
- Describe the two principles of inheritance proposed by Gregor Mendel.
- Explain how an individual’s genotype and phenotype differ.
- Explain the paired relationships among dominant and recessive alleles.
- Describe the structure and complementary base pairing of DNA.
- Describe natural selection and give an example of this occurring in a population.
- List evolutionary mechanisms that introduce new genetic variation into a population.
- Explain how divergent evolution and convergent evolution differ.
- Provide an example of how biotechnology is used in medicine.
- Explain how the scientific meaning of “theory” differs from the everyday meaning.
Applying concepts
- In pea plants, purple flowers ( P ) are dominant to white ( p ), and yellow peas ( Y ) are dominant to green ( y ). What are the possible genotypes and phenotypes for a cross between PpYY and ppYy pea plants? How many squares would you need in your table to complete an analysis of this cross?
- Galápagos medium ground finches are found on Santa Cruz and San Cristóbal islands, which are separated by about 100 kilometers of ocean. Occasionally, individuals from either island fly to the other island to stay. What evolutionary mechanism would alter the allele frequencies of the population? What might take place in this situation over time?
- Two species of fish had recently undergone sympatric speciation. The males of each species had a different coloring through which females could identify and choose a partner from her own species. After some time, pollution made the lake so cloudy it was hard for females to distinguish the colors of males. What might take place in this situation over time?
- Explain why the statement “a monkey is more evolved than a mouse” is incorrect.
